Выбор читателей





Популярные статьи
Вся система регуляции менструального цикла построена по иерархическому принципу (нижележащие структуры регулируются вышележащими, которые, в свою очередь, реагируют на изменения в нижележащих уровнях). При этом сигналы, поступающие от нижележащих структур, корректируют деятельность вышележащих. Репродуктивная система организована по иерархическому принципу. В ней выделяют пять уровней регуляции.
Первый уровень репродуктивной системы - экстрагипоталамические церебральные структуры. Они воспринимают импульсы из внешней среды и интерорецепторов и передают их через систему передатчиков нервных импульсов (нейротрансмиттеров) в нейросекреторные ядра гипоталамуса.
В регуляции функции репродуктивной системы участвует кора головного мозга. Поток информации, поступающий из внешнего мира, определяющий психическую деятельность, эмоциональный ответ и поведение, - все это сказывается на функциональном состоянии репродуктивной системы. Об этом свидетельствуют нарушения овуляции при острых и хронических стрессах, изменение менструального цикла при перемене климатических условий, ритма работы и т. д. Нарушения репродуктивной функции реализуются через изменение синтеза и потребления нейротрансмиттеров в нейронах мозга и в конечном счете через гипоталамические структуры ЦНС.
Второй уровень репродуктивной системы - гипофизотропная зона гипоталамуса. Над гипофизом в прямом и переносном смысле находится гипоталамус - структура головного мозга, регулирующая функционирование гипофиза. Гипоталамус состоит из скопления нервных клеток, часть которых продуцирует специальные гормоны (рилизинг-гормоны), оказывающие прямое действие на синтез гонадотропинов в гипофизе. В клетках гипоталамуса образуются гипофи-зотропные факторы (рилизинг-гормоны) - либерины. Рилизинг-гормон ЛГ (РГ-ЛГ люлиберин) и его синтетические аналоги обладают способностью стимулировать выделение ЛГ и ФСГ передней долей гипофиза.
Секреция РГ-ЛГ генетически запрограммирована и происходит в определенном пульсирующем ритме с частотой примерно один раз в час. Этот ритм получил название цирхорального (часового). Цирхоральный ритм выделения РГ-ЛГ формируется в пубертатном периоде и является показателем зрелости нейросекреторных структур гипоталамуса. Цирхоральная секреция РГ-ЛГ запускает гипоталамогипофизарно-яичниковую систему, но ее функцию нельзя считать автономной. Она моделируется импульсами из экстрагипоталамических структур.
Третий уровень репродуктивной системы - гипофиз, точнее, его передняя доля - аденогипофиз, в которой секретируются гонадотропные гормоны - фоллитропин (фолликулостимулирующий гормон, ФСГ), лютропин (лютеинизирующий гормон, ЛГ), пролактин (ПРЛ), регулирующие функции яичников и молочных желез.
Железой-мишенью ЛГ и ФСГ является яичник. ФСГ стимулирует рост фолликула, пролиферацию гранулезных клеток, индуцирует образование рецепторов ЛГ на поверхности клеток гранулезы. Под влиянием ФСГ увеличивается содержание ароматаз в зреющем фолликуле.
ЛГ стимулирует образование андрогенов (предшественников эстрогенов) в тека-клетках, совместно с ФСГ способствует овуляции и стимулирует синтез прогестерона в лютеинизированных клетках гранулезы овулировавшего фолликула.
Пролактин оказывает многообразное действие на организм женщины. Его основная биологическая роль - рост молочных желез и регуляция лактации. Он обладает также жиромобилизующим эффектом и оказывает гипотензивное действие. Увеличение секреции пролактина является одной из частых причин бесплодия, так как повышение его уровня в крови тормозит стероидогенез в яичниках и развитие фолликулов.
Четвертый уровень репродуктивной системы - яичники. В них происходят сложные процессы синтеза стероидов и развития фолликулов. Процесс фояликулогенеза происходит в яичнике непрерывно: начинается в антенатальном периоде и заканчивается в постменопауз альном.
Примордиальные фолликулы состоят из растущего ооцита, формирующейся прозрачной оболочки (zona pellucida) и нескольких слоев фолликулярного эпителия.
Дальнейший рост фолликула обусловлен превращением фолликулярного эпителия в многослойный, сек-ретирующий фолликулярную жидкость (liquor folliculi), которая содержит стероидные гормоны (эстрогены). Ооцит с окружающими его вторичной оболочкой и фолликулярными клетками, образующими лучистый венец (corona radiata) в виде яйценосного бугорка (cumulus oophoron), смещается к верхнему полюсу фолликула. Наружная оболочка дифференцируется на два слоя - внутренний и наружный. Вокруг разветвляющихся капилляров располагаются многочисленные интерстициальные клетки. Наружная оболочка фолликула (the-са folliculi externa) образована плотной соединительной тканью. Так выглядит вторичный фолликул (folliculi secundarii).
Зрелый фолликул, достигший своего максимального развития, заполненный фолликулярной жидкостью, называется третичным, или пузырчатым (folliculus ovaricus tertiams seu vesicularis). Он достигает таких размеров, что выпячивает поверхность яичника, причем яйценосный бугорок с ооцитом оказывается в выступающей части пузырька. Дальнейшее увеличение объема пузырька, переполненного фолликулярной жидкостью, приводит к растягиванию и разрыхлению как его наружной оболочки, так и белочной оболочки яичника в месте прилежания пузырька с последующим разрывом и овуляцией. Основная масса фолликулов (90%) претерпевает атретические изменения, и только очень небольшая часть их проходит полный цикл развития от примордиального фолликула, овулирует и превращается в желтое тело.
У приматов и человека в течение цикла развивается один фолликул. Доминантный фолликул уже в первые дни менструального цикла имеет диаметр 2 мм и в течение 14 дней, к моменту овуляции, увеличивается в среднем до 20-21 мм. В фолликулярной жидкости резко увеличивается содержание эстрадиола (Е2) и ФСГ. Подъем уровня эстрогенов (Е2) стимулирует выброс ЛГ и овуляцию.
Процесс овуляции представляет собой разрыв базальной мембраны доминантного фолликула и кровотечение из разрушенных капилляров, окружающих тека-клетки.
После выхода яйцеклетки в полость фолликула быстро врастают формирующиеся капилляры; гранулезные клетки подвергаются лютеинизации. Этот процесс ведет к образованию желтого тела, клетки которого секретируют прогестерон.
Желтое тело может быть менструальным (corpus luteum menstmationis), которое подвергается инволюции на 12- 14-й день, после чего образуется белое тело (corpus albicans), в дальнейшем исчезающее; или желтым телом беременности (corpus luteum graviditatis), которое образуется в случае оплодотворения и функционирует на протяжении всей беременности, достигая огромных размеров.
Материнской субстанцией для всех стероидных гормонов является холестерин, липопротеин низкой плотности, который поступает в яичник с током крови. Под влиянием ферментов происходят конечные этапы синтеза: превращение андрогенов в эстрогены.
В раннюю фолликулярную фазу менструального цикла в яичнике секретируется 60-100 мкг эстрадиола, в лютеиновую фазу - 270 мкг, к моменту овуляции - 400 - 900 мкг в сутки. Около 10% Е2 ароматизируется внегонадно из тестостерона. К моменту овуляции синтез эстрона возрастает до 600 мкг в сутки.
Прогестерона образуется в яичнике 2 мг/сут в фолликулярную фазу менструального цикла и 25 мг/сут - в лютеиновую фазу. В процессе метаболизма прогестерон в яичнике превращается в 20 альфа-дегидропрогестерон, обладающий сравнительно малой биологической активностью.
В яичнике синтезируется 1,5 мг/сут андростендиона, предшественника тестостерона. Столько же андростендиона образуется и в надпочечниках. Около 15% тестостерона под влиянием энзимов ароматизируется в дегидротестостерон - самый биологически активный андроген. Количество его в женском организме составляет 75 мкг/сутки.
Кроме того, в яичнике секретируются белковые вещества местного действия - окситоцин и релаксин. Окситоцин оказывает лютеолитическое действие, способствуя регрессу желтого тела. Релаксин оказывает токолитическое действие на миометрий и способствует овуляции. В яичниках образуются также простагландины.
Функцию репродуктивной системы, направленную на регуляцию овуляторного менструального цикла у женщин репродуктивного возраста, можно представить следующим образом.
В нейронах медиобазального гипоталамуса происходит пульсирующая секреция РГ-ЛГ в цирхоральном режиме. По аксонам нервных клеток нейросекрет (РГ-ЛГ) поступает в портальную систему и с кровью переносится в переднюю долю гипофиза.
Образование двух гонадотропинов (ЛГ и ФСГ) под влиянием одного РГ-ЛГ объясняется различной чувствительностью к нему клеток гипофиза, секретирующих ЛГ и ФСГ, а также различной скоростью их метаболизма. ФСГ и ЛГ гуморальным путем стимулируют рост фолликула, синтез стероидов и созревание яйцеклетки. Повышение уровня Е2 в преовуляторном фолликуле вызывает выброс ЛГ и ФСГ и овуляцию. Под влиянием ингибина тормозится выделение ФСГ. В клетках лютеинизированной гранулезы под влиянием ЛГ образуется прогестерон. Уменьшение содержания Е2 стимулирует выделение ЛГ и ФСГ.
Пятый уровень регуляции репродуктивной системы - ткани-мишени - точки приложения действия гормонов. Так называемые органы-мишени - органы, которые являются конечной точкой приложения половых гормонов, вырабатываемых яичниками. К ним относятся как органы репродуктивной системы (матка, маточные трубы, влагалище), так и другие органы (молочные железы, кожные покровы, кости, жировая ткань). Клетки названных тканей и органов содержат рецепторы к половым гормонам.
В головном мозге также обнаружены рецепторы к половым гормонам, что, по-видимому, может объяснять циклические колебания психики женщины в течение менструального цикла.
Итак, репродуктивная система представляет собой суперсистему, функциональное состояние которой определяется обратной афферентацией составляющих ее подсистем. Выделяют:
Обратная связь у половозрелой женщины имеет как отрицательный, так и положительный характер. Примером отрицательной связи является усиление выделения ЛГ передней долей гипофиза в ответ на низкий уровень эстрадиола в раннюю фолликулярную фазу цикла. Примером положительной обратной связи является выброс ЛГ и ФСГ в ответ на овуляторный максимум содержания эстрадиола в крови.
По механизму отрицательной обратной связи увеличивается образование РГ-ЛГ при снижении уровня ЛГ в клетках передней доли гипофиза. Примером ультракороткой отрицательной связи является увеличение секреции РГ-ЛГ при уменьшении его концентрации в нейросекреторных нейронах гипоталамуса.
В регуляции функции репродуктивной системы основными являются пульсирующая (цирхоральная) секреция РГ-ЛГ в нейронах гипоталамуса и регуляция выделения ЛГ и ФСГ эстрадиолом по механизму отрицательной и положительной обратной связи.
Л. Cycлoпapoвa
Женская репродуктивная система,
Список сокращений:
АДГ - антидиуретический гормон
АКТГ - кортиколиберин
аРГ-Гн - агонист рилизинг-гормона гонадотропинов
ЛГ - лютеинизирующий гормон
ОП - оксипрогестерон
РГ-Гн - рилизинг-гормон гонадотропинов
СТГ - соматолиберин
СЭФР - сосудистый эндотелиальный фактор роста
ТТГ - тиреотропный гормно (тиролиберин)
ФСГ - фолликулостимулирующий гормон
ФФР - фибропластический фактор роста
Нормальный менструальный цикл
Менструации - это кровянистые выделения из половых путей женщины, периодически воз-никающие в результате отторжения функцио-нального слоя эндометрия в конце двухфазного менструального цикла.
Комплекс циклических процессов, которые происходят в женском организме и внешне про-являются менструациями, называется менстру-альным циклом. Менструация начинается как реакция на изменение уровня стероидов, про-дуцируемых яичниками.
Клинические признаки нормального менструального циклаДлительность менструального цикла в активный репродуктивный период женщины составляет в среднем 28 дней. Продолжительность цикла от 21 до 35 дней считается нормальным. Большие промежутки наблюдаются во время полового со-зревания и климакса, что может быть проявлени-ем ановуляции, которая может отмечаться в это время наиболее часто.
Обычно менструация длится от 3 до 7 дней, количество теряемой крови незначительно. Уко-рочение или удлинение менструального кровоте-чения, а также появление скудных или обильных менструаций может служить проявлением ряда гинекологических заболеваний.
Характеристики нормального менструального цикла:
Длительность: 28±7 дней;
Длительность менструального кровотечения: 4±2 дня;
Объем кровопотери при менструации: 20-60 мл* ;
Средняя потеря железа: 16 мг
*
95 процентов здоровых женщин с каждой менструацией теряют менее 60 мл крови. Кровопотеря более 60-80 мл сочетается со снижением уровня гемоглобина, гематокрита и сывороточного железа.
Физиология менструального кровотечения:
Непосредственно перед менструацией развивается выраженный спазм спиральных артериол. После дилатации спиральных артериол начинается менструальное кровотечение. Поначалу адгезия тромбоцитов в сосудах эндометрия подавляется, но затем, по ме¬ре транссудации крови, поврежденные концы сосудов запечаты¬ваются внутрисосудистыми тромбами, состоящими из тромбо¬цитов и фибрина. Через 20 ч после начала менструации, когда большая часть эндометрия уже отторглась, развивается выражен¬ный спазм спиральных артериол, за счет чего и достигается гемо¬стаз. Регенерация эндометрия начинается через 36 ч после воз¬никновения менструации, несмотря на то, что отторжение эндо¬метрия еще полностью не закончено.
Регуляция менструального цикла является сложным нейрогуморальным механизмом, который осуществляется с участием 5 основных звеньев регуляции. К ним относятся: кора голов-ного мозга, подкорковые центры (гипоталамус), гипофиз, половые железы, периферические органы и ткани (матка, маточные трубы, влагалище, молочные железы, волосяные фолликулы, кости, жировая ткань). Последние носят название органов-мишеней, благодаря наличию рецепторов, чувствительных к действию гормонов, которые вырабатывает яичник на протяжении менструального цикла. Цитозолрецепторы - рецепторы цитоплазмы, обладают строгой специфичностью к эстрадиолу, прогестерону, тестостерону, в то время как ядерные рецепторы могут быть акцепторами таких молекул, как инсулин, глюкагон, аминопептиды.
Рецепторы к половым гормонам обнаруже-ны во всех структурах репродуктивной системы, а также в центральной нервной системе, коже, жировой и костной ткани и молочной железе. Свободная молекула стероидного гормона захватывается специфическим цитозолрецептором белковой природы, образующийся комплекс транслоцируется в ядро клетки. В ядре возникает новый комплекс с ядерным белковым рецептором; этот комплекс связывается с хроматином, регулирующим транскрипцию мРНК, участвующим в синтезе специфического тканевого белка. Внутриклеточный медиатор - циклическая аденозинмонофосфорная кислота (цАМФ) регулирует метаболизм в клетках ткани-мишени в соответствии с потребностями организма в ответ на воздействие гормонов. Основная масса стероидных гормонов (около 80% находится в крови и транспортируется в связанном виде. Транспорт их осуществляется специальными белками - стероидсвязывающими глобулинами и неспецифическими транспортными системами (альбуминами и эритроцитами). В связанном виде стероиды неактивны, поэтому глобулины, альбумины и эритроциты можно рассматривать как своеобразную буферную систему, контролирующую доступ стероидов к рецепторам клеток-мишеней.
Циклические функциональные изменения, происходящие в организме женщины, условно можно разделить на изменения в системе гипоталамус-гипофиз-яичники (яичниковый цикл) и матке, в первую очередь в ее слизистой оболочке (маточный цикл).
Наряду с этим, как правило, происходят циклические сдвиги во всех органах и системах женщины, в частности, в ЦНС, сердечно-сосудистой системе, системе терморегуляции, обменных процессах и т.д.
Гипоталамус
Гипоталамус - часть головного мозга, расположенная над зрительным перекрестом и образующая дно III желудочка. Это старый и стабильный компонент центральной нервной системы, общая организация которого мало менялась в процессе эволюции человека. Структурно и функционально гипоталамус связан с гипофизом. Выделяют три гипоталамические области: переднюю, заднюю и промежуточную. Каждая область образована ядрами — скоплениями тел нейронов определенного типа.
Помимо гипофиза гипоталамус влияет на лимбическую систему (миндалевидное тело, гиппокамп), таламус, мост. Указанные отделы также прямо или опосредованно влияют на гипоталамус.
Гипоталамус секретирует либерины и статины. Этот процесс регулируют гормоны, замыкающие три петли обратной связи: длинную, короткую и ультракороткую. Длинную петлю обратной связи обеспечивают циркулирующие половые гормоны, связывающиеся с соответствующими рецепторами в гипоталамусе, короткую: гормоны аденогипофиза, ультракороткую: либерины и статины. Либерины и статины регулируют активность аденогипофиза. Гонадолиберин стимулирует секрецию ЛГ и ФСГ, кортиколиберин - АКТГ, соматолиберин (СТГ), тиролиберин (ТТГ). Помимо либеринов и статинов в гипоталамусе синтезируются антидиуретический гормон и окситоцин. Эти гормоны транспортируются в нейрогипофиз, откуда попадают в кровь.
В отличие от капилляров других областей мозга капилляры воронки гипоталамуса фенестрированные. Именно они образуют первичную капиллярную сеть воротной системы.
В 70-80-х гг. была выполнена серия экспериментальных работ на обезьянах, которые позво-лили выявить различия функции нейросекреторных структур гипоталамуса приматов и грызунов. У приматов и человека аркуатные ядра медиобазального гипоталамуса являются единственным местом образования и выделения РГ-ЛГ, ответственного за гонадотропную функцию гипофиза. Секреция РГ-ЛГ генетически запрограммирована и происходит в определенном пульсирующем ритме с частотой примерно один раз в час. Этот ритм получил название цирхорального (часово-го). Область аркуатных ядер гипоталамуса по-лучила название аркуатного осциллятора. Цирхоральный характер секреции РГ-ЛГ был подтвержден путем прямого определения его в крови портальной системы ножки гипофиза и яремной вены у обезьян и в крови у женщин с овулятор-ным циклом.
Гормоны гипоталамуса
Рилизинг-гормон ЛГ выделен, синтезирован и подробно описан. Выделить и синтезировать фоллиберин до настоящего времени не удалось. РГ-ЛГ и его синтетические аналоги обладают способностью стимулировать выделение ЛГ и ФСГ передней долей гипофиза, поэтому в настоящее время принят один термин для гипоталамических гонадотропных либеринов - рилизинг-гормон гонадотропинов (РГ-Гн).
Гонадолиберин стимулирует секрецию ФСГ и ЛГ. Это декапептид, секретируемый нейронами ядра воронки. Гонадолиберин секретируется не постоянно, а в импульсном режиме. Он очень быстро разрушается протеазами (период полуразрушения составляет 2—4 мин), поэтому его импульсация должна быть регулярной. Частота и амплитуда выбросов гонадолиберина меняются на протяжении менструального цикла. Для фолликулярной фазы характерны частые колебания небольшой амплитуды уровня гонадолиберина в сыворотке крови. К концу фолликулярной фазы частота и ампли-туда колебаний возрастают, а затем снижаются на протяжении лютеиновой фазы.
Гипофиз
В гипофизе выделяют две доли: переднюю - аденогипофиз и заднюю - нейрогипофиз. Ней-рогипофиз имеет нейрогенное происхождение и представляет продолжение воронки гипоталамуса. Нейрогипофиз кровоснабжается из нижних гипофизарных артерий. Аденогипофиз развивается из эктодермы кармана Ратке, поэтому состоит из железистого эпителия и не имеет прямой связи с гипоталамусом. Синтезируемые в гипоталамусе либерины и статины попадают в аденогипофиз через особую воротную систему. Это основной источник кровоснабжения аденогипофиза. Кровь в воротную систему преимуще-ственно поступает через верхние гипофизарные артерии. В области воронки гипоталамуса они образуют первичную капиллярную сеть воротной системы, из нее формируются воротные вены, которые входят в аденогипофиз и дают начало вторичной капиллярной сети. Возможен обратный ток крови через воротную систему. Особенности кровоснабжения и отсутствие гематоэнцефалического барьера в области воронки гипоталамуса обеспечивают двустороннюю связь между гипоталамусом и гипофизом. В зависимости от окрашивания гематоксилином и эозином секреторные клетки аденогипофиза делят на хромофильные (ацидофильные) и базофильные (хромофобные). Ацидофильные клетки секретируют СТГ и пролактин, базофильные - ФСГ, ЛГ, ТТГ, АКТГ
Гормоны гипофиза
В аденогипофизе образуются СТГ, пролактин, ФСГ, ЛГ, ТТГ и АКТГ. ФСГ и ЛГ регулируют секрецию половых гормонов, ТТГ — секрецию тиреоидных гормонов, АКТГ — секрецию гормо-нов коры надпочечников. СТГ стимулирует рост, обладает анаболическим действием. Пролактин стимулирует рост молочных желез во время бе-ременности и лактацию после родов.
ЛГ и ФСГ синтезируются гонадотропными клетками аденогипофиза и играют важную роль в развитии яичниковых фолликулов. По структуре относятся к гликопротеинам. ФСГ стимулирует рост фолликула, пролиферацию гранулезных клеток, индуцирует образование рецепторов ЛГ на поверхности клеток гранулезы. Под влиянием ФСГ увеличивается содержание ароматаз в зреющем фолликуле. ЛГ стимулирует образование андрогенов (предшественников эстрогенов) в тека-клетках, совместно с ФСГ способствует овуляции и стимулирует синтез прогестерона в лютеинизированных клетках гранулезы овулировавшего фолликула.
Секреция ЛГ и ФСГ непостоянна и модулируется яичниковыми гормонами, особенно эстро-генами и прогестероном.
Таким образом, низкий уровень эстрогенов оказывает подавляющий эффект на ЛГ, в то вре-мя как высокий стимулирует его производство гипофизом. В поздней фолликулярной фазе уровень сывороточных эстрогенов достаточно высок, положительный эффект обратной связи утраивается, что способствует образованию преовуляторного пика ЛГ. И, наоборот, при терапии комбинированными контрацептивами уровень эстрогенов в сыворотке крови находится в пределах, определяющих отрицательную обратную связь, что приводит к снижению содержания гонадотропинов.
Механизм положительной обратной связи приводит к повышению в рецепторах концен-трации и продукции РГ-Гн.
В противоположность эффекту эстрогенов, низкий уровень прогестерона имеет положитель-ную реакцию обратной связи на секрецию ЛГ и ФСГ гипофизом. Такие состояния существуют непосредственно перед овуляцией и приводят к выбросу ФСГ. Высокий уровень прогестерона, который отмечается в лютеиновой фазе, уменьшает гипофизарную продукцию гонадотропинов. Малое количество прогестерона стимулирует вы-свобождение гонадотропинов на уровне гипофиза. Отрицательный эффект обратной связи прогестерона проявляется путем уменьшения производства РГ-Гн и снижения чувствительности к РГ-Гн на уровне гипофиза. Положительный эффект обратной связи прогестерона происходит на гипофиз и включает в себя повышенную чувствительность к РГ-Гн. Эстрогены и проге-стерон не являются единственными гормонами, воздействующими на секрецию гонадотропинов гипофизом. Таким же эффектом обладают гормоны ингибин и активин. Ингибин подавляет гипофизарную ФСГ секрецию, активин ее сти-мулирует.
Пролактин — это полипептид, состоящий из 198 аминокислотных остатков, синтезируемый лактотропными клетками аденогипофиза. Секреция пролактина контролируется дофамином. Он синтезируется в гипоталамусе и тормозит секрецию пролактина. Пролактин оказывает многообразное действие на организм женщины. Его основная биологическая роль — рост молочных желез и регуляция лактации. Он обладает также жиромобилизующим эффектом и оказывает гипотензивное действие. Увеличение секреции пролактина является одной из частых причин бесплодия, так как повышение его уровня в крови тормозит стероидогенез в яичниках и развитие фолликулов.
Окситоцин — пептид, состоящий из 9 аминокислотных остатков. Он образуется в нейро-нах крупноклеточной части паравентрикулярных ядер гипоталамуса. Основными мишенями окситоцина у человека служат гладкомышечные волокна матки и миоэпителиальные клетки молочных желез.
Антидиуретический гормон (АДГ) — это пептид, состоящий из 9 аминокислотных остатков. Синтезируется в нейронах супраоптического ядра гипоталамуса. Основная функция АДГ — регуляция ОЦК, АД, осмоляльности плазмы.
Яичниковый цикл
В яичниках проходит три фазы менструального цикла:
Фолликулярная фаза:
Одним из основных моментов фолликулярной фазы менструального цикла является развитие яйцеклетки. Яичник женщины представляет собой сложный орган, состоящий из многих компонентов, в результате взаимодействия которых секретируются половые стероидные гормоны и образуется готовая к оплодотворению яйцеклет-ка в ответ на циклическую секрецию гонадотро-пинов.
Стероидогенез
Гормональная активность от преантрального до периовуляторного фолликула описана как теория «две клетки, два гонадотропина». Стероидогенез происходит в двух клетках фолликула: в тека- и гранулезных клетках. В тека-клетках ЛГ стимулирует производство андрогенов из холестерола. В гранулезных клетках ФСГ стимулирует превращение полученных андрогенов в эстрогены (ароматизация). Дополнительно к эффекту ароматизации ФСГ так же отвечает за пролиферацию гранулезных клеток. Хотя известны другие медиаторы в развитии фолликулов яичника, эта теория является основной для по-нимания процессов, происходящих в фолликуле яичника. Выявлено, что для нормального цикла с достаточным уровнем эстрогенов необходимы оба гормона.
Производство андрогенов в фолликулах так же может регулировать развитие преантрально-го фолликула. Низкий уровень андрогенов усиливает процесс ароматизации, следовательно, увеличивает производство эстрогенов, и наоборот, высокий — тормозит процесс ароматизации и вызывает атрезию фолликула. Баланс ФСГ и ЛГ необходим для раннего развития фолликула. Оптимальным условием для начальной стадии развития фолликула является низкий уровень ЛГ и высокий ФСГ, что имеет место в начале мен-струального цикла. Если же уровень ЛГ высокий, тека-клетки производят большое количество андрогенов, вызывая атрезию фолликулов.
Выбор доминантного фолликула
Росту фолликула сопутствует секреция половых стероидных гормонов под влиянием ЛГ и ФСГ. Эти гонадотропины защищают группу преантральных фолликулов от атрезии. Одна-ко в норме только один из этих фолликулов развивается до преовуляторного, который затем освобождается и становится доминантным.
Доминантный фолликул в средней фолликулярной фазе является самым большим и наиболее развитым в яичнике. Уже в первые дни менструального цикла он имеет диаметр 2 мм и в течение 14 дней к моменту овуляции увеличивается в среднем до 21 мм. За это время происходит 100-кратное увеличение объема фолликулярной жидкости, количество выстилающих базальную мембрану клеток гранулезы увеличивается с 0,5х10 6 до 50х10 6 . Такой фолликул имеет самую высокую ароматизирующую активность и самую высокую концентрацию индуцированных ФСГ рецепторов к ЛГ, поэтому доминирующий фолликул выделяет самое высокое количество эстрадиола и ингибина. Далее ингибин усиливает синтез андрогенов под влиянием ЛГ, который является субстратом для синтеза эстрадиола.
В отличие от уровня ФСГ, который по мере увеличения концентрации эстрадиола снижа-ется, уровень ЛГ продолжает расти (в низких концентрациях эстрадиол тормозит секрецию ЛГ). Именно длительная эстрогенная стимуляция готовит овуляторный пик ЛГ. Одновремен-но с этим происходит подготовка доминантного фолликула к овуляции: под местным действием эстрогенов и ФСГ на гранулезных клетках увеличивается число рецепторов ЛГ. Выброс ЛГ приводит к овуляции, образованию желтого тела и увеличению секреции прогестерона. Овуляция происходит спустя 10—12 ч после пика ЛГ или спустя 32—35 ч после начала подъема его уровня. Обычно овулирует только один фолликул.
Во время выбора фолликула уровень ФСГ понижается в ответ на отрицательный эффект от эстрогенов, поэтому доминирующий фолликул является единственным, который продолжает свое развитие при падающем уровне ФСГ
Яичниково-гипофизарная связь является ре-шающей при выборе доминирующего фолликула и при развитии атрезии остальных фолликулов.
Ингибин и активин
Рост и развитие яйцеклетки, функционирование желтого тела происходит при взаимодей-ствии аутокринных и паракринных механизмов. Необходимо отметить два фолликулярных гор-мона, играющих значительную роль в стероидогенезе, — ингибин и активин.
Ингибин представляет собой пептидный гормон, вырабатываемый гранулезными клетками растущих фолликулов, снижает продукцию ФСГ. Кроме того, влияет на синтез андрогенов в яич-нике. Ингибин влияет на фолликулогенез следующим образом: уменьшая ФСГ до такого уровня, при котором развивается только доминантный фолликул.
Активин представляет собой пептидный гормон, вырабатывается в гранулезных клетках фолликулов и гипофиза. По данным некоторых авторов, активин вырабатывается также и плацентой. Активин увеличивает производство ФСГ гипофизом, усиливает процесс связывания ФСГ с гранулезными клетками.
Инсулиноподобные факторы роста
Инсулиноподобные факторы роста (ИФР-1 и ИФР-2) синтезируются в печени под влиянием гормона роста и, возможно, в гранулезных клетках фолликулов, действуют как паракрин-ные регуляторы. Перед овуляцией содержание ИФР-1 и ИФР-2 в фолликулярной жидкости повышается за счет увеличения количества самой жидкости в доминантном фолликуле. ИФР-1 участвует в процессе синтеза эстрадиола. ИФР-2 (эпидермальный) тормозит синтез стероидов в яичниках.
Овуляция:
Овуляторный пик ЛГ приводит к повыше-нию концентрации простагландинов и активности протеаз в фолликуле. Сам процесс овуляции представляет собой разрыв базальной мембра-ны доминантного фолликула и кровотечение из разрушенных капилляров, окружающих тека-клетки. Изменения в стенке преовуляторного фолликула, обеспечивающие ее истончение и разрыв, происходят под влиянием фермента коллагеназы; определенную роль играют также простагландины, содержащиеся в фолликулярной жидкости, протеолитические ферменты, образующиеся в гранулезных клетках, окситопин и релаксин. В результате этого в стенке фолликула образуется небольшое отверстие, через которое медленно выходит яйцеклетка. Непосредственные измерения показали, что давление внутри фолликула во время овуляции не возрастает.
В конце фолликулярной фазы ФСГ воздей-ствует на рецепторы ЛГ в гранулезных клетках. Эстрогены являются обязательным кофактором в этом эффекте. По мере развития доминантного фолликула продукция эстрогенов увеличивается. В итоге производство эстрогенов достаточно для достижения секреции гипофизом ЛГ, что приводит к увеличению его уровня. Повышение происходит вначале очень медленно (с 8-го по 12-й день цикла), затем быстро (после 12-го дня цикла). В течение этого времени ЛГ активирует лютеинизацию гранулезных клеток в доминантном фолликуле. Таким образом, происходит выделение прогестерона. Далее прогестерон усиливает эффект эстрогенов на секрецию гипофизом ЛГ, приводя к повышению его уровня.
Овуляция происходит в течение 36 ч после начала подъема ЛГ. Определение выброса ЛГ является одним из лучших методов, который определяет овуляцию и проводится с помощью прибора «определитель овуляции».
Периовуляторный пик ФСГ, вероятно, происходит в результате положительного эффекта прогестерона. В дополнение к повышению содержания ЛГ, ФСГ и эстрогенов, во время ову-ляции отмечается и повышение уровня сывороточных андрогенов. Эти андрогены выделя-ются в результате стимулирующего эффекта ЛГ на тека-клетки, особенно в недоминирующем фолликуле.
Увеличение содержания андрогенов оказывает воздействие на усиление либидо, подтверждая, что этот период у женщин наиболее фертильный.
Уровень ЛГ стимулирует мейоз, после того как сперматозоид попадает в яйцеклетку. При выделении ооцита из яичника в овуляцию происходит разрушение стенки фолликула. Это ре-гулируется ЛГ, ФСГ и прогестероном, которые стимулируют активность протеолитических эн-зимов, таких как плазминогеновые активаторы (которые выделяют плазмин, стимулирующий активность коллагеназы) и простагландины. Простагландины не только увеличивают активность протеолитических энзимов, но так же способствуют появлению воспалительно-подобной реакции в стенке фолликула и стимулируют ак-тивность гладкой мускулатуры, что способствует выходу ооцита.
Важность простагландинов в процессе овуля-ции доказана исследованиями, которые определяют, что снижение выделения простагландина может привести к задержке освобождения ооцита из яичника при нормальном стероидогенезе (синдром неразвивающегося лютеинизированного фолликула— СНЛФ). Так как СНЛФ нередко является причиной бесплодия, женщинам, желающим забеременеть, рекомендуется избе-гать приема синтезированных ингибиторов простагландина.
Лютеиновая фаза:
Строение желтого тела
После выброса яйцеклетки из яичника в полость фолликула быстро врастают формирующиеся капилляры; гранулезные клетки подвергаются лютеинизации: увеличению в них цитоплазмы и образо-ванию липидных включений. Гранулезные клетки и текоциты образуют желтое тело — основной регулятор лютеиновой фазы менструального цикла. Клетки, образовавшие стенку фолликула, накапливают липиды и желтый пигмент лютеин и начинают секретировать прогестерон, эстрадиол-2, ингибин. Мощная сосудистая сеть спо-собствует поступлению гормонов желтого тела в системный кровоток. Полноценное желтое тело развивается только в тех случаях, когда в преовуляторном фолликуле образуется адекватное число гранулезных клеток с высоким содержанием рецепторов ЛГ. Увеличение размеров желтого тела после овуляции происходит в основном за счет увеличения размеров гранулезных клеток, в то время как число их не увеличивается из-за отсутствия митозов. У человека желтое тело секретирует не только прогестерон, но и эстрадиол и андрогены. Механизмы регрессии желтого тела изучены недостаточно. Известно, что лютеолитическим действием обладают простагландины.

Рис. Ультразвуковая картина "цветущего" желтого тела при беременности 6 нед. 4 дня. Режим энергетического картирования.
Гормональная регуляция лютеиновой фазы
Если беременность не наступает, происходит инволюция желтого тела. Этот процесс регулируется по механизму отрицательной обратной связи: гормоны (прогестерон и эстрадиол), секретируемые желтым телом, действуют на гонадотропные клетки гипофиза, подавляя секрецию ФСГ и ЛГ. Секрецию ФСГ подавляет также ингибин. Снижение уровня ФСГ, а также местное действие прогестерона препятствует развитию группы примордиальных фолликулов.
Существование желтого тела зависит от уровня секреции ЛГ. При его снижении, обычно через 12-16 дней после овуляции, происходит инволюция желтого тела. На его месте образуется белое тело. Механизм инволю-ции неизвестен. Скорее всего, она обусловлена паракринными влияниями. По мере инволюции желтого тела уровень эстрогенов и прогестерона падает, что приводит к повышению секреции гонадотропных гормонов. По мере повышения содержания ФСГ и ЛГ начинает развиваться новая группа фолликулов.
Если произошло оплодотворение, существование желтого тела и секрецию прогестерона поддерживает хорионический гонадотропин. Таким образом, имплантация эмбриона приводит к гормональным изменениям, которые сохраняют желтое тело.
Длительность лютеиновой фазы у большинства женщин постоянна и составляет примерно 14 дней.
Гормоны яичников
Сложный процесс биосинтеза стероидов за-вершается образованием эстрадиола, тестосте-рона и прогестерона. Стероидпродуцирующими тканями яичников являются клетки гранулезы, выстилающие полость фолликула, клетки внутренней теки и в значительно меньшей степени строма. Клетки гранулезы и тека-клетки синергично участвуют в синтезе эстрогенов, клетки текальной оболочки являются главным источником андрогенов, которые в незначительном количестве образуются и в строме; прогестерон синтезируется в тека-клетках и клетках гранулезы.
В яичнике в раннюю фолликулярную фазу менструального цикла секретируется 60-100 мкг эстрадиола (Е2), в лютеиновую фазу - 270 мкг, к моменту овуляции - 400-900 мкг в сутки. Около 10% Е2 ароматизируется в яичнике из тестостерона. Количество эстрона, образующегося в раннюю фолликулярную фазу, составляет 60-100 мкг, к моменту овуляции синтез его возрастает до 600 мкг в сутки. Только половина количества эстрона образуется в яичнике. Вторая половина ароматизируется в Е2. Эстриол является малоактивным метаболитом эстрадиола и эстрона.
Прогестерон образуется в яичнике в количестве 2 мг/сут в фолликулярную фазу и 25 мг/сут в лютеиновую фазу менструального цикла. В процессе метаболизма прогестерон в яичнике превращается в 20-дегидропрогестерон, обладающий сравнительно малой биологической активностью.
В яичнике синтезируются следующие андрогены: андростендион (предшественник те-стостерона) в количестве 1,5 мг/сут (столько же андростендиона образуется в надпочечниках). Из андростендиона образуется около 0,15 мг те-стостерона, примерно такое же количество его образуется в надпочечниках.
Краткий обзор процессов, происходящих в яичниках
Фолликулярная фаза:
ЛГ стимулирует продукцию андрогенов в тека-клетках.
ФСГ стимулирует продукцию эстрогенов в гранулезных клетках.
Наиболее развитый фолликул в середине фолликулярной фазы становится доминантным.
Увеличивающееся образование эстрогенов и ингибина в доминантном фолликуле, по-давляет выделение ФСГ гипофизом.
Снижение уровня ФСГ вызывает атрезию всех фолликулов кроме доминантного.
Овуляция:
ФСГ индуцирует ЛГ рецепторы.
Протеолитические ферменты в фолликуле приводят к разрушению его стенки и осво-бождению ооцита.
Лютеиновая фаза:
Желтое тело образуется из гранулезных и тека-клеток, сохранившихся после овуляции.
Прогестерон, выделяемый желтым телом, является доминирующим гормоном. При отсутствии беременности лютеолизис происходит через 14 дней после овуляции.
Маточный цикл
Эндометрий состоит из двух слоев: функционального и базального. Функциональный слой меняет свою структуру под действием половых гормонов и, если беременность не наступила, отторгается во время менструации.
Пролиферативная фаза:
Началом менструального цикла считают 1-й день менструации. По окончании менструации толщина эндометрия составляет 1-2 мм. Эндометрий состоит практически только из базального слоя. Железы узкие, прямые и короткие, выстланы низким цилиндрическим эпителием, цитоплазма клеток стромы почти не различает-ся. По мере увеличения уровня эстрадиола формируется функциональный слой: эндометрий готовится к имплантации эмбриона. Железы удлиняются и становятся извитыми. Возрастает число митозов. По мере пролиферации высота эпителиальных клеток возрастает, а сам эпителий из однорядного становится многорядным к моменту овуляции. Строма отечна и разрыхлена, в ней увеличиваются ядра клеток и объем цитоплазмы. Сосуды умеренно извиты.
Секреторная фаза:
В норме овуляция происходит на 14-й день менструального цикла. Секреторная фаза ха-рактеризуется высоким уровнем эстрогенов и прогестерона. Однако после овуляции число ре-цепторов эстрогенов в клетках эндометрия снижается. Пролиферация эндометрия постепенно тормозится, снижается синтез ДНК, уменьшается число митозов. Таким образом, преимущественное влияние на эндометрий в секреторную фазу оказывает прогестерон.
В железах эндометрия появляются содержа-щие гликоген вакуоли, которые выявляются с помощью ШИК-реакции. На 16-й день цикла эти вакуоли достаточно крупные, имеются во всех клетках и находятся под ядрами. На 17-й день ядра, оттесненные вакуолями, располагаются в центральной части клетки. На 18-й день вакуоли оказываются в апикальной части, а ядра — в базальной части клеток, гликоген начинает выделяться в просвет желез путем апокриновой се-креции. Наилучшие условия для имплантации создаются на 6—7-й день после овуляции, т.е. на 20—21-й день цикла, когда секреторная активность желез максимальна.
На 21-й день цикла начинается децидуальная реакция стромы эндометрия. Спиральные артерии резко извиты, позднее вследствие уменьше-ния отека стромы, они видны отчетливо. Сна-чала появляются децидуальные клетки, которые постепенно образуют скопления. На 24-й день цикла эти скопления формируют периваскулярные эозинофильные муфты. На 25-й день образуются островки децидуальных клеток. К 26-му дню цикла децидуальная реакция становится число нейтрофилов, которые мигрируют туда из крови. Нейтрофильная инфильтрация сменяется некрозом функционального слоя эндометрия.
Менструация:
Если имплантация не произошла, железы перестают продуцировать секрет, а в функцио-нальном слое эндометрия начинаются дегенеративные изменения. Непосредственная причина его отторжения — резкий спад содержания эстрадиола и прогестерона в результате инволюции желтого тела. В эндометрии снижается венозный отток и происходит расширение сосудов. Далее происходит сужение артерий, что приводит к ишемии и повреждению тканей и функциональной потере эндометрия. Затем происходит кровотечение из фрагментов артериол, оставшихся в базальном слое эндометрия. Мен-струация прекращается при сужении артерий, эндометрий восстанавливается. Таким образом, прекращение кровотечения в сосудах эндометрия отличается от гемостаза в других частях организма.
Как правило, кровотечение прекращается в результате скопления тромбоцитов и отложения фибрина, что приводит к рубцеванию. В эндометрии рубцевание может привести к потере его функциональной активности (синдром Ашермана). Для избежания этих последствий необходима альтернативная система гемостаза. Сокращение сосудов является механизмом остановки кровотечения в эндометрии. При этом рубцевание минимизировано фибринолизом, который разрушает кровяные сгустки. Позже восстановление эндометрия и образование новых кровеносных сосудов (ангиогенез) приводит к завершению кровотечения в течение 5-7 дней от начала менструального цикла.
Влияние отмены эстрогенов и прогестерона на менструацию определено четко, но остается неясной роль паракринных медиаторов. Вазоконстрикторы: простагландин F2a, эндотелии-1 и тромбоцит-активизирующий фактор (ТАФ) могут производиться в пределах эндометрия и участвовать в сокращении сосудов. Так же они способствуют началу менструации и дальнейшему контролю над ней. Эти медиаторы могут регулироваться воздействием сосудорасширяющих агентов, таких как простагландин Е2, простациклин, оксид азота, которые вырабатываются эндометрием. Простагландин F2a обладает выраженным сосудосуживающим действием, усиливает спазм артерий и ишемию эндометрия, вызывает сокращения миометрия, что, с одной стороны, уменьшает кровоток, с другой - способствует удалению отторгнутого эндометрия.
Восстановление эндометрия включает в себя железистую и стромальную регенерацию и ангиогенез. Сосудистый эндотелиальный фактор роста (СЭФР) и фибропластический фактор ро-ста (ФФР) обнаружены в эндометрии и являются сильными ангиогенезивными агентами. Выявлено, что эстрогенпродуцированная железистая и стромальная регенерация усиливается под воздействием эпидермальных факторов роста (ЭФР). Такие факторы роста как трансформирующий фактор роста (ТФР) и интерлейкины, особенно интерлейкин-1 (ИЛ-1), имеют большое значение.
Краткий обзор процессов, происходящих в эндометрии
Менструация:
Основную роль в начале менструации играет спазм артериол.
Функциональный слой эндометрия (верхний, составляющий 75% толщины) отторгается.
Менструация прекращается вследствие спазма сосудов и восстановления эндометрия. Фибринолиз препятствует образованию спаек.
Пролиферативная фаза:
Характеризуется индуцированной эстроге-нами пролиферацией желез и стромы.
Секреторная фаза:
Характеризуется индуцированной прогестероном секрецией желез.
В позднюю секреторную фазу индуцируется децидуализация.
Децидуализация представляет собой необратимый процесс. При отсутствии наступления беременности в эндометрии происходит апоптоз с последующим появлением менструации.
Итак, репродуктивная система представляет собой суперсистему, функциональное состоя-ние которой определяется обратной афферентацией составляющих ее подсистем. Выделяют: длинную петлю обратной связи между гормонами яичника и ядрами гипоталамуса; между гормонами яичника и гипофизом; короткую петлю между передней долей гипофиза и гипоталамусом; ультракороткую между РГ-ЛГ и нейроцитами (нервными клетками) гипоталамуса.
Обратная связь у половозрелой женщины имеет как отрицательный, так и положительный характер. Примером отрицательной связи является усиление выделения ЛГ передней долей гипофиза в ответ на низкий уровень эстрадиола в раннюю фолликулярную фазу цикла. Примером положительной обратной связи является выброс ЛГ и ФСГ в ответ на овуляторный максимум содержания эстрадиола в крови. По механизму отрицательной обратной связи увеличивается образование РГ-ЛГ при снижении уровня ЛГ в клетках передней доли гипофиза.
Резюме
Гонадолиберин синтезируется нейронами ядра воронки, затем попадает в воротную систему гипофиза и поступает по ней в аде-ногипофиз. Секреция гонадолиберина про-исходит импульсивно.
Ранний этап развития группы примордиальных фолликулов не зависит от ФСГ.
По мере инволюции желтого тела снижается секреция прогестерона и ингибина и повышается уровень ФСГ.
ФСГ стимулирует рост и развитие группы примордиальных фолликулов и секрецию ими эстрогенов.
Эстрогены готовят матку к имплантации, стимулируя пролиферацию и дифференцировку функционального слоя эндометрия и вместе с ФСГ способствуют развитию фолликулов.
Согласно двухклеточной теории синтеза половых гормонов, ЛГ стимулирует синтез андрогенов в текоцитах, которые затем под влиянием ФСГ превращаются в эстрогены в гранулезных клетках.
Рост концентрации эстрадиола по механизму отрицательной обратной связи, петля
которой замыкается в гипофизе и гипоталамусе, подавляет секрецию ФСГ.
Фолликул, который будет овулировать в данном менструальном цикле, называет-ся доминантным. В отличие от остальных фолликулов, начавших рост, он несет большее число рецепторов ФСГ и синтезирует большее количество эстрогенов. Это позво-ляет ему развиваться, несмотря на снижение уровня ФСГ.
Достаточная эстрогенная стимуляция обе-спечивает овуляторный пик ЛГ. Он, в свою очередь, вызывает овуляцию, образование желтого тела и секрецию прогестерона.
Функционирование желтого тела зависит от уровня ЛГ. При его снижении желтое тело подвергается инволюции. Обычно это происходит на 12—16-й день после овуляции.
Если произошло оплодотворение, существование желтого тела поддерживает хорионический гонадотропин. Желтое тело продолжает секретировать прогестерон, необходимый для сохранения беременно-сти на ранних сроках.
Лекция для врачей "Роль гормонов в регуляции менструального цикла". Курс лекций по патологическому акушерству для студентов медицинского колледжа. Лекцию для врачей проводит
Дьякова С.М, врач акушер-гинеколог, преподаватель общий стаж работы 47 лет.
Роль гормонов в регуляции менструального цикла. Часть 1.
Роль гормонов в регуляции менструального цикла. Часть 2.
Роль гормонов в регуляции менструального цикла. Часть 3.
Менструальный цикл и его регуляция
Репродуктивная система (РС) выполняет множество функций, наиболее важной из которых является продолжение биологического вида. Оптимальной функциональной активности репродуктивная система достигает к 16-18 годам, когда организм готов к зачатию, вынашиванию и вскармливанию ребенка. Особенностью РС является также постепенное угасание различных функций: к 45 годам угасает генеративная, к 50 - менструальная, затем - гормональная функции.
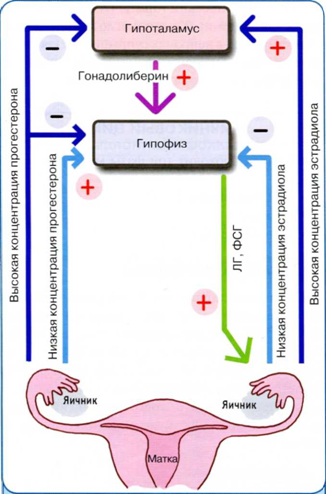
Репродуктивная система состоит из пяти уровней: экстрагипаталамического (кора головного мозга), гипоталамуса, гипофиза, яичников и органов и тканей- мишеней (рис. 1).
Репродуктивная система работает по иерархическому принципу, т.е. нижележащий уровень подчиняется вышележащему (за счет прямых связей между звеньями регуляции). Основой регуляции функций РС является принцип отрицательной обратной связи между различными уровнями (рис. 1), т.е. при снижении концентрации периферических гормонов (яичниковых, в частности, эстрадиола), усиливаются синтез и выделение гормонов гипоталамуса и гипофиза (гонадотропин-рилизинг гормона (ГиРГ) и гонадотропных гормонов соответственно). Особенностью регуляции женской РС является и наличие положительной обратной связи, когда в ответ на значительное повышение уровня эстрадиола в преовуляторном фолликуле увеличивается продукция ГнРГ и гонадотропинов (овуляторный пик выделения ЛГ и ФСГ). Функционирование репродуктивная система женщины характеризуется цикличностью (повторяемостью) процессов регуляции, представления о которых укладываются в современное понятие менструального цикла.
Менструальный цикл - это повторяющиеся изменения в деятельности системы гипоталамус-гипофиз-яичники и вызванные ими структурные и функциональные изменения репродуктивных органов: матки, маточных труб, молочных желез, влагалища.
Кульминацией каждого цикла является менструальное кровотечение (менструация), первый день которого считается началом менструального цикла. Первая в жизни девочки менструация называется менархе, средний возраст менархе - 12-14 лет.
Рис. 1. Регуляция женской репродуктивной системы: РГ – рилизинг-гормоны, ФСГ– фолликулостимулирующий гормон, ЛГ – лютеинизирующий гормон, ТТГ- тиреотропный гормон, АКТГ – адренокортикотропный гормон, Прл – пролактин, Т4 – тироксин, АДГ – антидиуретический гормон, А – андрогены, Э – эстрогены, П– прогестерон, И – ингибин, Р – факторы роста; сплошные стрелки – прямые связи, пунктирные стрелки – обратные отрицательные связи.
Продолжительность менструального цикла определяется от первого дня одной до первого дня следующей менструации и составляет в норме от 21 до 35 дней (у подростков в течение 1,5-2 лет после менархе продолжительность цикла может быть более вариабельной – от 21 до 40-45 дней). Такой цикл называется нормопонирующим. Разновидностью нормопонирующего цикла является идеальный цикл продолжительностью 28 дней. Укорочение менструального цикла (менее 21 дня) называется антепонацией (антепонирующий цикл), удлинение (более 35 дней) – постпонацией (постпонирующий цикл).
Продолжительность нормальной менструации составляет в среднем 3-5 дней (в норме – от 3 до 7 дней), а средняя кровопотеря – 50-70 мл (в норме – до 80 мл).
Менструальный цикл условно подразделяют на яичниковый и маточный циклы. Яичниковый (овариальный) цикл подразумевает циклические процессы, происходящие в яичниках под воздействием гонадотропных и рилизинг-гормонов. Циклические изменения в организме женщины носят двухфазный характер. Первая (фолликулиновая, фолликулярная) фаза цикла определяется созреванием фолликула и яйцеклетки в яичнике, после чего происходят его разрыв и выход из него яйцеклетки – овуляция. Вторая (лютеиновая) фаза связана с образованием желтого тела. Одновременно в циклическом режиме в эндометрии последовательно происходят регенерация и пролиферация функционального слоя, сменяющаяся секреторной активностью его желез, заканчивающиеся десквамацией функционального слоя (менструация). Циклические процессы в эндометрии представляют собой последовательно сменяющие друг друга фазы маточного цикла.
Биологическое значение изменений, которые происходят на протяжении менструального цикла в яичниках и эндометрии, состоит в обеспечении репродуктивной функции на этапах созревания яйцеклетки, ее оплодотворения и имплантации зародыша в матке. Если оплодотворения яйцеклетки не происходит, функциональный слой эндометрия отторгается, из половых путей появляются кровянистые выделения, а в репродуктивной системе вновь и в той же последовательности происходят процессы, направленные на обеспечение созревания яйцеклетки.
Высшим V -м уровнем регуляции менструального цикла является кора головного мозга , а именно лимбическая система и амигдалоидные ядра. Кора головного мозга осуществляет контроль над гипаталамо-гипофизарной системой посредством нейромедиаторов (нейротрансмиттеров), т.е. передатчиков нервного импульса на нейросекреторные ядра гипоталамуса. Наиболее важная роль отводится нейропептидам (дофамину, норадреналину, серотонину, кисс-пептину, семейству опиоидных пептидов), а также гормону эпифиза мелатонину. При стрессовых ситуациях, при перемене климата, ритма работы (например, ночные смены) наблюдаются нарушения овуляции, реализующиеся через изменение синтеза и потребления нейротрансмиттеров в нейронах мозга, а также мелатонина в эпифизе.
В ЦНС имеется большое количество рецепторов к эстрадиолу и другим стероидным гормонам, что указывает на их важную роль не только в реализации обратных связей, но и в нейромедиаторном обмене.
IV уровень репродуктивной системы – гипоталамус – представляет собой высший вегетативный центр, гибрид нервной и эндокринной систем, координирующий функции всех внутренних органов и систем, поддерживающий гомеостаз в организме. Под контролем гипоталамуса находится гипофиз и регуляция эндокринных желез: гонад (яичников), щитовидной железы, надпочечников (рис. 1). В гипоталамусе имеется два типа нейросекреторных клеток, осуществляющих гипоталамо-гипофизарное взаимодействие:
Местом синтеза гонадотропного рилизинг-гормона (ГнРГ) являются аркуатные ядра медиобазального гипоталамуса. Выделен, синтезирован и описан рилизинг-гормон к ЛГ – люлиберин. Выделить и синтезировать фоллилиберин до настоящего времени не удалось. Поэтому гипоталамические гонадотропные либерины обозначают ГнРГ, так как они стимулируют выделение как ЛГ, так и ФСГ передней долей гипофиза. Секреция ГнРГ генетически запрограммирована и происходит в определенном пульсирующем ритме – 1 раз в 60-90 мин (цирхоральный, часовой, ритм секреции). В настоящее время доказана пермиссивная (запускающая) роль ГнРГ в функционировании РС. Пульсовой ритм секреции ГнРГ формируется в пубертатном возрасте и является показателем зрелости нейросекретных структур гипоталамуса. Цирхоральная секреция ГнРГ запускает гипоталамо- гипофизарно-яичниковую систему. Под влиянием ГнРГ происходит выделение ЛГ и ФСГ из передней доли гипофиза.
Секреция ГнРГ модулируется нейропептидами экстрагипоталамических структур, а также половыми гормонами по принципу обратной связи. В ответ на повышение преовуляторного пика эстрадиола повышается синтез и выделение ГнРГ, под влиянием которого усиливается секреция гонадотропинов, в результате чего происходит овуляция. Прогестерон оказывает и ингибирующий, и стимулирующий эффект на продукцию гонадотропинов, действуя по принципу обратной связи как на уровне гипоталамуса, так и на уровне гипофиза (рис. 1).
Основная роль в регуляции выделения пролактина принадлежит дофаминэргическим структурам гипоталамуса. Дофамин (ДА) тормозит выделение пролактина из гипофиза, тиреолиберин – стимулирует. Антагонисты дофамина усиливают выделение пролактина.
Нейросекреты гипоталамуса оказывает биологическое действие на организм различными путями. Основной путь – парагипофизарный через вены, впадающие в синусы твердой мозговой оболочки, а оттуда в системный кровоток. Трансгипофизарный путь – через систему воротной (портальной) вены к передней доле гипофиза; особенностью портальной кровеносной системы является возможность тока крови в ней в обе стороны (как к гипоталамусу, так и к гипофизу), что важно для реализации механизмов обратных связей. Обратное влияние на гипофиз половых гормонов яичников осуществляется через вертебральные артерии.
Таким образом, циклическая секреция ГнРГ запускает гипоталамо-гипофизарно-яичниковую систему, но ее функцию нельзя считать автономной, она модулируется как нейропептидами ЦНС, так и яичниковыми стероидами по принципу обратной связи.
III уровень – передняя доля гипофиза (аденогипофиз). В аденогипофизе различают три вида клеток хромофобные (резервные), ацидофильные и базофильные. Здесь синтезируются гонадотропные гормноны: фолликулостимулирующий гормон, или фоллитропин (ФСГ), лютеинизирующий, или лютеотропин (ЛГ); а также пролактин (Прл) и другие тропные гормоны: тиреотропный гормон, тиреотропин (ТТГ), соматотропный гормон (СТГ), адренокортикотропный гормон, кортикотропин (АКТГ); меланостимулирующий гормон, меланотропин (МСГ) и липотропный (ЛПГ) гормон. ЛГ и ФСГ являются гликопротеидами, Прл – полипептидом.
Секреция ЛГ и ФСГ контролируется (рис. 1):
ЛГ и ФСГ определяют первые этапы синтеза половых стероидов в яичниках путем взаимодействия со специфическими рецепторами в тканях гонад. Эффективность гормональной регуляции определяется как количеством активного гормона, так и уровнем содержания рецепторов в клетке-мишени.
Пролактин (Прл) – полипептид, синтезируемый клетками аденогипофиза (лактотрофами), контролирует лактацию, стимулирует рост протоков молочных желез, поддерживает функцию желтого тела и синтез прогестерона, обладает различными биологическими эффектами: снижает минеральную плотность костной ткани, повышает активность клеток поджелудочной железы, приводя к инсулинорезистентности (диабетогенное действие), участвует в регуляции обмена веществ, пищевого поведения, циклов сна и бодрствования, либидо и др.
II уровень репродуктивной системы – яичники. Основной структурной единицей яичника является фолликул, содержащий яйцеклетку (ооцит). В половых железах происходит рост и созревание фолликулов, овуляция, образование желтого тела, синтез половых стероидов.
Процесс фолликулогенеза в яичниках происходит непрерывно – с антенатального периода до постменопаузы. При рождении в яичниках девочки находится примерно 2 млн. примордиальных (первичных зародышевых) фолликулов. Основная их масса претерпевает атретические изменения (атрезия – обратное развитие) в течение всей жизни и только очень небольшая часть проходит полный цикл развития от примордиального до зрелого с овуляцией и образованием в последующем желтого тела. Ко времени менархе в яичниках содержится 200-450 тыс. примордиальных фолликулов (так называемый овариальный резерв). Из них в течение жизни могут овулировать только 400-500, остальные подвергаются атрезии (около 90%). В процессе атрезии фолликулов важная роль отводится апоптозу (программируемой клеточной гибели) – биологическому процессу, в результате которого происходит полное рассасывание клетки под влиянием собственного лизосомального аппарата. На протяжении одного менструального цикла развивается, как правило, только один фолликул с яйцеклеткой внутри. В случае созревания большего числа возможна многоплодная беременность.
Важная роль в механизмах ауто- и паракринной регуляции функции не только овариальной, но и всей репродуктивной системы принадлежит факторам роста.
Факторы роста (ФР) – биологически активные вещества, стимулирующие или ингибирующие дифференцировку клеток, передающих гормональный сигнал. Они синтезируются в неспецифических клетках различных тканей организма и обладают аутокринным, паракринным, интракринным и эндокринным эффектом. Аутокринный эффект реализуется путем воздействия на клетки, непосредственно синтезирующие данный ФР. Паракринный – реализуется действием на соседние клетки. Интракринный эффект – ФР действует как внутриклеточный мессенджер (передатчик сигнала). Эндокринный эффект реализуется через кровоток на отдаленные клетки.
Наиболее важную роль в физиологии репродуктивной системы играют следующие ФР: инсулиноподобные (ИФР), эпидермальный (ЭФР), трансформирующие (ТФР- α, ТФР-β), сосудистый эндотелиальный (васкулоэндотелиальный) фактор роста (СЭФР), ингибины, активины, антимюллеров гормон (АМГ).
Инсулиноподобные факторы роста I и II (ИФР-I , ИФР-II ) синтезируются в гранулезных клетках и других тканях, стимулируют синтез андрогенов в тека-клетках яичника, ароматизацию андрогенов в эстрогены, пролиферацию клеток гранулезы, образование рецепторов к ЛГ на гранулезных клетках. Их продукция регулируется инсулином.
Эпидермальный фактор роста (ЭФР) – наиболее сильный стимулятор клеточной пролиферации, обнаруживается в клетках гранулезы, строме эндометрия, молочных железах и других тканях; обладает онкогенным эффектом в эстрогензависимых тканях (эндометрий, молочные железы).
Сосудистый эндотелиальный фактор роста (СЭФР) играет важную роль в ангиогенезе растущих фолликулов, а также мио- и эндометрия. СЭФР повышает митогенную активность эндотелиальных клеток, проницаемость сосудистой стенки. Экспрессия СЭФР повышена при эндометриозе, миоме матки, опухолях яичников и молочных желез, СПКЯ и др.
Трансформирующие факторы роста (ТФР-α , ТФР-β ) стимулируют клеточную пролиферацию, участвуют росте и созревании фолликулов, пролиферации клеток гранулезы; оказывают митогенный и онкогенный эффект, экспрессия их повышена при раке эндометрия, яичников. К белковым веществам семейства ТФР-β относят ингибины, активин, фоллистатин, а также АМГ.
Ингибины (А и В) – белковые вещества, образуются в клетках гранулезы и других тканях, участвуют в регуляции синтеза ФСГ, тормозя ее, подобно эстрадиолу, по сходному механизму обратной связи. Образование ингибина В возрастает в середине фолликулярной фазы цикла параллельно повышению концентраций эстрадиола после выбора доминантного фолликула, а достигнув максимума, тормозит выделение ФСГ.
Активин обнаружен в гранулезных клетках фолликула и гонадотрофах гипофиза, стимулирует синтез ФСГ, пролиферацию клеток гранулезы, ароматизацию андрогенов в эстрогены, подавляет синтез андрогенов в тека- клетках, предотвращает спонтанную (преждевременную, до овуляции) лютеинизацию преовуляторного фолликула, стимулирует продукцию прогестерона в желтом теле.
Фоллистатин – ФСГ-блокирующий белок, секретируется клетками передней доли гипофиза, гранулезы; подавляет секрецию ФСГ.
Антимюллеров гормон (АМГ) – представитель семейства ТФР-β, продуцируется у женщин в гранулезных клетках преантральных и малых антральных фолликулов, играет важную роль в механизмах рекрутирования и селекции фолликулов, является количественным показателем овариального резерва и используется в клинической практике для его оценки и прогнозирования ответа яичников на стимуляцию овуляции, а также может служить маркером гранулезоклеточных опухолей яичников, при которых АМГ существенно повышается. АМГ не контролируется гонадотропинами, не вовлечен в классическую петлю обратной связи (в отличие от ФСГ, эстрадиола и ингибина В), не зависит от фазы цикла и действует как паракринный фактор регуляции репродуктивной системы.
В яичнике женщины фолликулы находятся на различных стадиях зрелости. Фолликулогенез начинается с 12-й недели антенатального развития; основная масса фолликулов подвергается атрезии. До конца не известно, какие факторы ответственны за рост примордиальных фолликулов. Примордиальные фолликулы характеризуются одним слоем плоских прегранулезных клеток, небольшим незрелым ооцитом (не завершившим второе деление мейоза), клетки тека (оболочки) отсутствуют.

Рис. 2. Стадии развития фолликула

Таким образом, общая продолжительность фолликулогенеза от момента инициации роста примордиальных фолликулов до овуляции зрелого фолликула составляет порядка 200 дней; на фолликулярную фазу очередного менструального цикла приходится лишь завершающая стадия формирования доминантного фолликула и овуляция. Поскольку процессы фолликулогенеза происходят непрерывно, этим можно объяснить наличие в яичниках фолликулов различных стадий зрелости, определяемых эхографически, в любой день менструального цикла (рис. 3).
Овариальный цикл состоит из двух фаз: фолликулярной и лютеиновой. Отсчет фолликулярной фазы цикла начинается с первого дня очередной менструации, при идеальном менструальном цикле первая фаза продолжается около 2-х недель, характеризуется ростом и созреванием доминантного фолликула и завершается его овуляцией, происходящей на 13- 14-й дни цикла. Затем наступает лютеиновая фаза цикла, продолжающаяся с 14-15-го по 28-й дни, в течение которой происходит формирование, развитие и регресс желтого тела. При антепонирующем или постпонирующем цикле продолжительность фолликулярной фазы может отличаться от таковой в идеальном или близком к нему цикле.
Гонадотропин-зависимый рост фолликулов начинается в конце предыдущего менструального цикла. Повышение синтеза и выделения ФСГ гипофизом происходит по принципу отрицательной обратной связи в ответ на снижение уровня прогестерона, эстрадиола и ингибина В при регрессе желтого тела. Под действием ФСГ продолжается рост антральных фолликулов и в ранней фолликулярной фазе менструального цикла (4-5-й дни от начала менструации) их размеры составляют 4-5 мм в диаметре. В этот период ФСГ стимулирует пролиферацию и дифференцировку клеток гранулезы, синтез в них ЛГ-рецепторов, активацию ароматаз и синтез эстрогенов и ингибина. ЛГ в ранней фолликулярной фазе влияет преимущественно на синтез андрогенов – предшественников эстрогенов.
Максимального значения ФСГ достигает к 5-6-му дню менструального цикла, после чего снижается (под действием увеличивающихся концентраций эстрадиола и ингибина В, синтезируемых гранулезой растущих антральных фолликулов), затем опять повышается одновременно с ЛГ к овуляторному пику на 13-14-й дни цикла (рис. 4). Селекция доминантного фолликула происходит к 5-7-му дню цикла из пула антральных фолликулов диаметром 5-10 мм. Доминантным становится фолликул с наибольшим диаметром, с наибольшим числом клеток гранулезы и рецепторов к ФСГ, благодаря чему доминантный фолликул сохраняет способность к дальнейшему росту и синтезу эстрадиола несмотря на снижение уровня ФСГ в крови. Дальнейший рост доминантного фолликула, начиная с середины фолликулярной фазы цикла, становится не только ФСГ-зависимым, но и ЛГ- и ФСГ-зависимым. В быстром росте лидирующего фолликула играют роль и возрастающие концентрации эстрадиола и ФР – ИФР, СЭФР. К моменту овуляции доминантный фолликул достигает размеров 18-21 мм (рис. 3). В остальных антральных фолликулах снижение сывороточного уровня ФСГ вызывает процессы атрезии (апоптоза). В механизмах атрезии незрелых фолликулов определенная роль отводится высоким концентрациям андрогенов, синтезирующихся в этих же маленьких фолликулах (рис. 2, 3).
Овуляция – разрыв зрелого фолликула и выход из него яйцеклетки. Процесс овуляции происходит при достижении максимального уровня эстрадиола в преовуляторном фолликуле (рис. 4), который по положительной обратной связи стимулирует овуляторный выброс ЛГ и ФСГ гипофизом. Овуляция происходит через 10-12 ч. после пика ЛГ или через 24-36 ч. после пика эстрадиола (рис. 4). Процесс разрыва базальной мембраны фолликулапроисходит подвлиянием различных ферментов и биологически активных веществ в лютеинизированных клетках гранулезы: протеолитических ферментов, плазмина, гистамина, коллагеназы, простагландинов, окситоцина и релаксина. Показана важная роль прогестерона, который синтезируется в лютеинизированных клетках преовуляторного фолликула под влиянием пика ЛГ, в активации протеолитических ферментов, участвующих в разрыве базальной мембраны фолликула. Овуляция сопровождается кровотечением из разрушенных капилляров, окружающих тека-клетки.

После овуляции в полость овулировавшего фолликула быстро врастают образующиеся капилляры, клетки гранулезы подвергаются дальнейшей лютеинизации с образованием желтого тела, секретирующего прогестерон под влиянием ЛГ. Лютеинизация гранулезных клеток морфологически проявляется в увеличении их объема и образовании липидных включений. Желтое тело - транзиторное гормонально-активное образование, функционирующее в течение 14 дней независимо от общей продолжительности менструального цикла. Полноценное желтое тело развивается только в фазе, когда в преовуляторном фолликуле образуется адекватное количество гранулезных клеток с высоким содержанием рецепторов ЛГ. В развитии желтого тела различают следующие стадии:
Если происходит зачатие и имплантация плодного яйца (на 21-22 дни цикла), формирующийся хорион начинает продуцировать человеческий хорионический гонатропин (ХГЧ), который стимулирует дальнейшее развитие желтого тела. В этом случае формируется желтое тело беременности, которое продолжает синтезировать прогестерон в больших концентрациях, необходимых для пролонгирования беременности. Желтое тело беременности существует до 8-10 недели гестации, затем подвергается регрессу, а гормональную поддержку беременности берет на себя сформировавшаяся к концу 1-го триместра плацента.
Циклические процессы в яичнике характеризуются не только морфологическими изменениями фолликулов и желтого тела, но и неразрывно связанными с ними процессами стероидогенеза – образования половых гормонов. В настоящее время общепринятой считается двуклеточная теория биосинтеза стероидов в яичниках, согласно которой ЛГ стимулирует синтех андрогенов в тека-клетках, тогда как ФСГ – синтез ферментов ароматаз, метаболизирующих андрогены в эстрогены в клетках гранулезы.
Стероидпродуцирующими структурами яичников являются клетки гранулезы, тека и, в меньшей степени, строма. Тека-клетки являются главным источником андрогенов, клетки гранулезы – эстрогенов, прогестерон синтезируется в тека-клетках и максимально в лютеиновых клетках желтого тела (лютеинизированных клетках гранулезы). Субстратом для всех стероидов, в том числе надпочечниковых и тестикулярных, является холестерин (рис. 5).

Синтез половых гормонов происходит также и внегонадно. Известно, что в жировой ткани имеется энзимная система Р450 ароматаза, которая участвует в превращении андрогенов в эстрогены. Этот процесс может быть инициирован различными митогенными ФР или самим эстрадиолом. Кроме того, биологически активный тестостерон (дигидротестостерон) также синтезируется внегонадно в периферических тканях-мишенях (волосяные фолликулы, сальные железы) под влиянием фермента 5-α-редуктазы.
Около 96% всех половых стероидов находится в связанном с белками состоянии, в частности, с глобулином связывающим половые стероиды (ГСПС), а также альбуминами, синтез которых осуществляется в печени. Биологическое действие гормонов определяется несвязанными, свободными фракциями, уровень которых изменяется при различных патологических состояниях, в частности инсулинорезистентности, заболеваниях печени и др.
Эстрогены. Основными фракциями эстрогенов являются эстрон (Е 1 ), эстрадиол (Е 2 ), эстриол (Е 3 ). Наиболее биологически активным является эстрадиол. Эстриол явялется периферическим метаболитом эстрона и эстрадиола, а не самостоятельным продуктом секреции яичников. В 1965 г. был описан и четвертый эстроген – эстетрол (Е 4 ), до настоящего времени мало изученный, обладающий слабым эстрогенным действием.
Биологическое действие эстрогенов на различные органы и ткани зависит от числа и вида специфических рецепторов и их чувствительности. Установлено наличие двух типов рецепторов к эстрадиолу: ЭР- α – ядерные рецепторы, оказывающие пролиферативное действие, и мембранные ЭР- β , оказывающие антипролиферативное действие.
Гестагены. Основным гестагеном является прогестерон, образующийся преимущественно в желтом теле яичников.
Андрогены. Основными фракциями андрогенов являются сильный андроген тестостерон, его слабый предшественник андростендион, а также дигидроандростендион (ДГЭА) и его сульфат (ДГЭА-С). Наиболее биологически активным является метаболит тестостерона – дигидротестостерон, синтезирующийся в периферических тканях-мишенях (волосяные фолликулы, сальные железы) под влиянием фермента 5-α- редуктазы. Основными местами синтеза андрогенов в женском организме являются яичники, надпочечники, а также жировая ткань и кожа с ее придатками.
Циклические изменения в эндометрии касаются его функционального поверхностного слоя, состоящего из компактных эпителиальных клеток, и промежуточного, которые отторгаются во время менструации. Базальный слой, не отторгаемый во время менструации, обеспечивает восстановление десквамированных слоев.
Циклические превращения функционального слоя эндометрия протекают соответственно яичниковому циклу в три последовательные стадии– стадия пролиферации, стадия секрецииистадиядесквамации (менструация).
Фаза десквамации. Наблюдающиеся в конце каждого менструального цикла менструальное кровотечение обусловлено отторжением функционального слоя эндометрия. Начало менструации считается первым днем менструального цикла. Продолжительность менструального кровотечения в среднем составляет 3-5 дней. В связи с регрессом желтого тела и резким снижением содержания половых стероидов в эндометрии нарастает гипоксия. Началу менструации способствует длительный спазм артерий, приводящий к стазу крови и образованию тромбов. Гипоксию тканей (тканевой ацидоз) усугубляют повышенная проницаемость эндотелия, ломкость стенок сосудов, многочисленные мелкие кровоизлияния и массивная лейкоцитарная инфильтрация. Выделяемые из лейкоцитов лизосомальные протеолитические ферменты усиливают расплавление тканевых элементов. Вслед за длительным спазмом сосудов наступает их паретическое расширение с усиленным притоком крови. При этом отмечаются рост гидростатического давления в микроциркуляторном русле и разрыв стенок сосудов, которые к этому времени в значительной степени утрачивают свою механическую прочность. На этом фоне и происходит активная десквамация некротизированных участков функционального слоя. К концу 1-х суток менструации отторгается 2/3 функционального слоя, а полная его десквамация обычно заканчивается на 3-й день.
Менструальные выделения содержат кровь и шеечную слизь, богаты лейкоцитами. Менструальная кровь почти не свертывается, она богата ионами кальция, содержит мало фибриногена и лишена протромбина. В среднем за менструацию женщина теряет 50 – 70 мл крови.
Сразу же после отторжения некротизированного эндометрия наступает стадия регенерации , характеризующаяся эпителизацией раневой поверхности эндометрия за счет клеток базального слоя. Процессы регенерации происходят под контролем эстрогенов и способствуют, наряду со спазмом сосудов и тромбообразованием, остановке менструального кровотечения. Некоторые авторы выделяют регенерацию в отдельную стадию маточного цикла.
Фаза пролиферации. Десквамация и регенерация слизистой после менструации заканчивается к 3-5-му дню цикла. Затем под действием увеличивающейся концентрации эстрогенов толщина функционального слоя увеличивается за счет роста всех элементов базального слоя: желез, стромы, кровеносных сосудов. Железы эндометрия имеют вид прямых или несколько извитых трубочек с прямым просветом. Спиральные артерии мало извиты. В стадии поздней пролиферации (11-14 день цикла) железы эндометрия становятся извитыми, штопорообразно закругляются, просвет их несколько расширен. Спиралевидные артерии, растущие из базального слоя достигают поверхности эндометрия, они несколько извиты. Толщина функционального слоя эндометрия к концу фазы пролиферации достигает 7-8 мм.
Фаза секреции (секреторной трансформации) начинается после овуляции на 13-14-й дни цикла, длится 14 дней и непосредственно связана с активностью желтого тела. Она характеризуется тем, что эпителий желез под влиянием прогестерона и эстрадиола начинает вырабатывать секрет, содержащий кислые гликозаминогликаны, гликопротеиды, гликоген.
В ранней стадии фазы секреции (15-18-й дни цикла) появляются первые признаки секреторных превращений. Железы становятся более извитыми, просвет их несколько расширен. В поверхностных слоях эндометрия могут быть очаговые кровоизлияния, связанные с кратковременным снижением эстрогенов после овуляции.
В средней стадии фазы секреции (19-23-й дни цикла), когда концентрация прогестерона максимальна и повышается уровень эстрогенов, функциональный слой эндометрия становится более высоким (9-12 мм) и отчетливо разделяется на 2 слоя. Глубокий (губчатый, спонгиозный) слой, граничащий с базальным, содержит большое количество сильно извитых желез и небольшое количество стромы. Плотный (компактный) слой составляет – 1/4-1/5 толщины функционального слоя. В нем меньше желез и больше соединительнотканных клеток. Наиболее выражена секреция на 20-21 дни цикла. К этому времени в строме эндометрия возникают децидуоподобные превращения (клетки компактного слоя становятся крупными, округлой или полигональной формы, в их цитоплазме появляется гликоген). Спиральные артерии резко извиты, образуют «клубки» и обнаруживаются во всем функциональном слое, проницаемость сосудов возрастает, просветы сосудов расширяются, объем кровоснабжения эндометрия возрастает. Эти изменения в железах и сосудах эндометрия составляют суть его преимплантационной подготовки и по времени синхронизированы с поступлением в полость матки плодного яйца (так называемое имплантационное окно – 7-е сутки после зачатия). Если имплантация пройдет успешно, в дальнейшем под действием увеличивающейся концентрации прогестерона эндометрий подвергнется децидуальной трансформации. При отсутствии беременности в эндометрии наступают дегенеративные изменения.
Поздняя стадия фазы секреции (24-27-й дни цикла) характеризуется нарушением трофики эндометрия и постепенным нарастанием в нем дегенеративных изменений. Уменьшается высота эндометрия, сморщивается строма функционального слоя, усиливается складчатость стенок желез, и они приобретают звездчатые или пилообразные очертания. На 26-27-й день цикла в поверхностных слоях компактного слоя наблюдаются лакунарное расширение капилляров и очаговые кровоизлияния в строму. Состояние эндометрия, подготовленного таким образом к распаду и отторжению, называется анатомической менструацией и обнаруживается за сутки до начала клинической менструации (кровотечения).
Слизистая оболочка перешейка матки по морфологическому строению сходна с эндометрием, однако в ней не различают функционального и базального слоя.
В канале шейки матки также происходят циклические изменения. Во время менструаций происходит десквамация не слизистой оболочки канала шейки матки, а лишь поверхностного ее эпителия. Под действием эстрогенов в фолликулярной фазе цикла цервикальный канал расширяется, наружный зев приоткрывается (положительный «симптом зрачка»), увеличивается продукция шеечной слизи, достигая максимума к моменту овуляции (положительный «симптом папоротника», «симптом натяжения шеечной слизи» – 8-10 см). Под влиянием прогестерона в лютеиновой фазе цикла цервикальныйканалсужается,наружныйзевзамыкается (отрицательный
«симптом зрачка»), шеечная слизь становится густой, плотной, не растягивается (табл. 1), слизистая оболочка шейки матки, влагалища приобретает цианотичный оттенок.
Циклические изменения происходят и в слизистой оболочке влагалища , которая представлена многослойным плоским неороговевающим эпителием.Так в первую половину цикла, под влиянием эстрогенов
происходит пролиферация промежуточного и поверхностного слоев слизистой оболочки. Во влагалищном мазке преобладают зрелые, поверхностные клетки, карио-пикнотический индекс (КПИ) высокий – 60-80% в преовуляторный период (табл. 1). Во вторую фазу цикла под влиянием прогестерона идет апоптоз и слущивание поверхностных клеток. В мазке преобладают промежуточные клетки, они принимают вытянутую форму и располагаются преимущественно группами (индекс скученности; КПИ низкий – 20-25%, см. табл. 1).
Таблица 1. Тесты функциональной диагностики

Примечание: ТФД – тесты функциональной диагностики, КПИ – карио- пикнотический индекс, БТ – базальная температура; дни менструального цикла: 0 – день овуляции, цифры со знаком «-» – дни до овуляции (фолликулярная фаза цикла), цифры со знаком «+» – дни после овуляции (лютеиновая фаза цикла).
В молочных железах под влиянием эстрогенов в первой половине менструального цикла идет пролиферация эпителия млечных ходов, а во вторую фазу под влиянием прогестерона – пролиферация секреторного эпителия в ацинусах (дольках).
Посмотреть и купить книги по УЗИ Медведева:
УЗИ Анализы
Менструальный цикл женщины – это сложный процесс, который осуществляется циклически в организме женщины под воздействием специальных веществ-гормонов, способствующих созреванию яйцеклетки в яичниках один раз в месяц.
Гормоны – это биологически активные химические вещества, регулирующие деятельность органов и организма в целом. Гормоны вырабатываются эндокринными железами (щитовидная железа, надпочечники, гипофиз, гипоталамус, половые железы и др.).
Половые гормоны вырабатываются в яичниках у женщин и в яичках у мужчин. Половые гормоны бывают женские и мужские. Мужские половые гормоны-андрогены, к ним относится и тестостерон. К женским гормонам относятся эстрогены и прогестерон.
В организме женщины присутствуют не только женские гормоны, но и небольшое количество мужских половых гормонов-андрогенов. Половые гормоны играют главную роль в регуляции менструального цикла. В каждом менструальном цикле организм женщины готовится к беременности. Менструальный цикл можно разделить на несколько периодов (фаз).
1 фаза
(фолликулярная или развитие яйцеклеток). В эту фазу внутренняя оболочка матки (эндометрий) отторгается и начинается менструация. Сокращения матки в этот период могут сопровождаться болями внизу живота. У некоторых женщин менструация короткая - 2 дня, у других длится 7 дней. В первой половине менструального цикла в яичниках растёт фолликул, где развивается и созревает яйцеклетка, которая затем выходит из яичника (овуляция). Эта фаза длится от 7 до 21 дней, что зависит от многих факторов.
Овуляция обычно происходит с 7 по 21 день цикла, чаще в середине месячного цикла (примерно на 14 день). Покинув яичник, созревшая яйцеклетка продвигается к матке по маточным трубам.
2 фаза (образование желтого тела). После овуляции лопнувший фолликул преобразуется в жёлтое тело, которое вырабатывает гормон прогестерон. Это главный гормон, поддерживающий беременность . В это время в матке происходит процесс подготовки к принятию оплодотворённой яйцеклетки. Внутренняя оболочка матки (эндометрит) утолщается и обогащается питательными веществами. Обычно эта фаза около 14 дней после овуляции. Если оплодотворение не произошло, наступит менструация.
Начало менструального цикла первый день менструации, поэтому длительность цикла определяется с первого для менструации до первого дня следующей менструации. В норме продолжительность менструального цикла колеблется от 21 до 35 дня. Средняя продолжительность цикла составляет 28 дней.
ППри встрече яйцеклетки со сперматозоидом происходит оплодотворение. Оплодотворенная яйцеклетка прикрепляется к стенке матки и образуется плодное яйцо. Начинают вырабатываться в большом объеме гормоны, "выключающие" менструальный цикл на весь срок беременности. Под действием этих же гормонов происходит функциональное изменение в организме женщины, подготовка ее к родам.
1 - маточная труба; 2 - дно матки; 3 - воронка; 4 - фимбрии; 5 - фимбриальный отдел маточной трубы; 6 - желтое тело; 7 - слизистая оболочка матки; 8 - яичник; 9 - связка яичника; 10 - полость матки; 11 - внутренний зев; 12 - канал шейки матки; 13 - наружный зев; 14 - влагалище.
Здравствуйте. У меня проблема задержки месячных (их нет почти месяц). Я, конечно, понимаю, что нужно идти к врачу… Но не могли бы вы подсказать, мог ли сбой быть из-за гастрита, который обнаружился совсем недавно, и двух отравлений подряд?
Ответ: Здравствуйте. Если Вы исключили возможность беременности, то сопутствующие заболевания могут явиться причиной задержки менструации.
Глава 2. НЕЙРОЭНДОКРИННАЯ РЕГУЛЯЦИЯ МЕНСТРУАЛЬНОГО ЦИКЛА
Менструальный цикл - генетически детерминированные, циклически повторяющиеся изменения в организме женщины, особенно в звеньях репродуктивной системы, клиническим проявлением которых служат кровяные выделения из половых путей (менструация).
Менструальный цикл устанавливается после менархе (первой менструации) и сохраняется в течение репродуктивного (детородного) периода жизни женщины до менопаузы (последней менструации). Циклические изменения в организме женщины направлены на возможность воспроизводства потомства и носят двухфазный характер: 1-я (фолликулярная) фаза цикла определяется ростом и созреванием фолликула и яйцеклетки в яичнике, после чего происходят разрыв фолликула и выход из него яйцеклетки - овуляция; 2-я (лютеиновая) фаза связана с образованием желтого тела. Одновременно в циклическом режиме происходят последовательные изменения в эндометрии: регенерация и пролиферация функционального слоя, сменяющаяся секреторной трансформацией желез. Изменения в эндометрии заканчиваются десквамацией функционального слоя (менструацией).
Биологическое значение изменений, которые происходят на протяжении менструального цикла в яичниках и эндометрии, состоит в обеспечении репродуктивной функции после созревания яйцеклетки, ее оплодотворения и имплантации зародыша в матке. Если оплодотворения яйцеклетки не происходит, функциональный слой эндометрия отторгается, из половых путей появляются кровяные выделения, а в репродуктивной системе вновь и в той же последовательности происходят процессы, направленные на обеспечение созревания яйцеклетки.
Менструация - это повторяющиеся с определенными интервалами кровяные выделения из половых путей на протяжении всего репродуктивного периода, исключая беременность и лактацию. Менструация начинается в конце лютеиновой фазы менструального цикла в результате отторжения функционального слоя эндометрия. Первая менструация (menarhe) возникает в возрасте 10-12 лет. В течение последующих 1-1,5 лет менструации могут быть нерегулярными, и лишь затем устанавливается регулярный менструальный цикл.
Первый день менструации условно принимается за 1-й день менструального цикла, а продолжительность цикла рассчитывается как интервал между первыми днями двух следующих одна за другой менструаций.
Внешние параметры нормального менструального цикла:
Длительность - от 21 до 35 дней (у 60% женщин средняя продолжительность цикла составляет 28 дней);
Продолжительность менструальных выделений - от 3 до 7 дней;
Величина кровопотери в менструальные дни - 40-60 мл (в среднем
50 мл).
Процессы, обеспечивающие нормальное течение менструального цикла, регулируются единой функционально-связанной нейроэндокринной системой, включающей центральные (интегрирующие) отделы, периферические (эффекторные) структуры, а также промежуточные звенья.
Функционирование репродуктивной системы обеспечивается строго генетически запрограммированным взаимодействием пяти основных уровней, каждый из которых регулируется вышележащими структурами по принципу прямой и обратной, положительной и отрицательной взаимосвязи (рис. 2.1).
Первым (высшим) уровнем регуляции репродуктивной системы являются кора головного мозга и экстрагипоталамические церебральные структуры
(лимбическая система, гиппокамп, миндалевидное тело). Адекватное состояние ЦНС обеспечивает нормальное функционирование всех нижележащих звеньев репродуктивной системы. Различные органические и функциональные изменения в коре и подкорковых структурах могут приводить к нарушениям менструального цикла. Хорошо известна возможность прекращения менструаций при сильных стрессах (потеря близких людей, условия военного времени и т.д.) или без очевидных внешних воздействий при общей психической неуравновешенности ("ложная беременность" - задержка менструации при сильном желании беременности или, наоборот, при ее боязни).
Специфические нейроны головного мозга получают информацию о состоянии как внешней, так и внутренней среды. Внутреннее воздействие осуществляется с помощью специфических рецепторов к стероидным гормонам яичников (эстрогенам, прогестерону, андрогенам), находящимся в ЦНС. В ответ на воздействие факторов внешней среды на кору головного мозга и экстрагипоталамические структуры происходят синтез, выделение и метаболизм нейротрансмиттеров и нейропептидов. В свою очередь, ней-ротрансмиттеры и нейропептиды влияют на синтез и выделение гормонов нейросекреторными ядрами гипоталамуса.
К важнейшим нейротрансмиттерам, т.е. веществам-передатчикам нервных импульсов, относятся норадреналин, дофамин, γ-аминомасляная кислота (ГАМК), ацетилхолин, серотонин и мелатонин. Норадреналин, аце-тилхолин и ГАМК стимулируют выброс гонадотропного рилизинг-гормона (ГнРГ) гипоталамусом. Дофамин и серотонин уменьшают частоту и снижают амплитуду выработки ГнРГ в течение менструального цикла.
Нейропептиды (эндогенные опиоидные пептиды, нейропептид Y, га-ланин) также участвуют в регуляции функции репродуктивной системы. Опиоидные пептиды (эндорфины, энкефалины, динорфины), связываясь с опиатными рецепторами, приводят к подавлению синтеза ГнРГ в гипоталамусе.
Рис. 2.1. Гормональная регуляция в системе гипоталамус - гипофиз - периферические эндокринные железы - органы мишени (схема): РГ - рилизинг-гормоны; ТТГ - тиреотропный гормон; АКТГ - адренокотикотропный гормон; ФСГ - фол-ликулостимулирующий гормон; ЛГ - лютеинизирующий гормон; Прл - пролак-тин; П - прогестерон; Э - эстрогены; А - андрогены; Р - релаксин; И - инги-бин; Т 4 - тироксин, АДГ - антидиуретический гормон (вазопрессин)
Вторым уровнем регуляции репродуктивной функции является гипоталамус. Несмотря на малые размеры, гипоталамус участвует в регуляции полового поведения, осуществляет контроль за вегетососудистыми реакциями, температурой тела и другими жизненно важными функциями организма.
Гипофизотропная зона гипоталамуса представлена группами нейронов, составляющих нейросекреторные ядра: вентромедиальное, дорсомедиаль-ное, аркуатное, супраоптическое, паравентрикулярное. Эти клетки имеют свойства как нейронов (воспроизводящие электрические импульсы), так и эндокринных клеток, вырабатывающих специфические нейросекреты с диаметрально противоположными эффектами (либерины и статины). Ли-берины, или рилизинг-факторы, стимулируют освобождение соответствующих тропных гормонов в передней доле гипофиза. Статины оказывают ин-гибирующее действие на их выделение. В настоящее время известны семь либеринов, по своей природе являющихся декапептидами: тиреолиберин, кортиколиберин, соматолиберин, меланолиберин, фоллиберин, люлибе-рин, пролактолиберин, а также три статина: меланостатин, соматостатин, пролактостатин, или пролактинингибирующий фактор.
Люлиберин, или рилизинг-гормон лютеинизирующего гормона (РГЛГ), выделен, синтезирован и подробно описан. Выделить и синтезировать фол-ликулостимулирующий рилизинг-гормон до настоящего времени не удалось. Однако установлено, что РГЛГ и его синтетические аналоги стимулируют выделение гонадотрофами не только ЛГ, но и ФСГ. В связи с этим принят один термин для гонадотропных либеринов - "гонадотропин-рилизинг-гормон" (ГнРГ), по сути, являющийся синонимом люлиберина (РГЛГ).
Основное место секреции ГнРГ - аркуатные, супраоптические и пара-вентрикулярные ядра гипоталамуса. Аркуатные ядра воспроизводят секреторный сигнал с частотой приблизительно 1 импульс в 1-3 ч, т.е. в пульсирующем или цирхоральном режиме (circhoral - вокруг часа). Эти импульсы имеют определенную амплитуду и вызывают периодическое поступление ГнРГ через портальную систему кровотока к клеткам аденогипофиза. В зависимости от частоты и амплитуды импульсов ГнРГ в аденогипофизе происходит преимущественная секреция ЛГ или ФСГ, что, в свою очередь, вызывает морфологические и секреторные изменения в яичниках.
Гипоталамо-гипофизарная область имеет особую сосудистую сеть, которая называется портальной системой. Особенностью данной сосудистой сети является возможность передачи информации как от гипоталамуса к гипофизу, так и обратно (от гипофиза к гипоталамусу).
Регуляция выделения пролактина в большей степени находится под ста-тиновым влиянием. Дофамин, образующийся в гипоталамусе, тормозит высвобождение пролактина из лактотрофов аденогипофиза. Увеличению секреции пролактина способствуют тиреолиберин, а также серотонин и эндогенные опиоидные пептиды.
Кроме либеринов и статинов, в гипоталамусе (супраоптическом и пара-вентрикулярном ядрах) продуцируются два гормона: окситоцин и вазопрес-син (антидиуретический гормон). Гранулы, содержащие данные гормоны, мигрируют от гипоталамуса по аксонам крупноклеточных нейронов и накапливаются в задней доле гипофиза (нейрогипофиз).
Третьим уровнем регуляции репродуктивной функции является гипофиз, он состоит из передней, задней и промежуточной (средней) доли. Непосредственное отношение к регуляции репродуктивной функции имеет передняя доля (аденогипофиз) . Под воздействием гипоталамуса в аденогипофизе се-кретируются гонадотропные гормоны - ФСГ (или фоллитропин), ЛГ (или лютропин), пролактин (Прл), АКТГ, соматотропный (СТГ) и тиреотропный (ТТГ) гормоны. Нормальное функционирование репродуктивной системы возможно лишь при сбалансированном выделении каждого из них.
Гонадотропные гормоны (ФСГ, ЛГ) передней доли гипофиза находятся под контролем ГнРГ, который стимулирует их секрецию и высвобождение в кровеносное русло. Пульсирующий характер секреции ФСГ, ЛГ является следствием "прямых сигналов" из гипоталамуса. Частота и амплитуда импульсов секреции ГнРГ меняется в зависимости от фаз менструального цикла и влияет на концентрацию и соотношение ФСГ/ЛГ в плазме крови.
ФСГ стимулирует в яичнике рост фолликулов и созревание яйцеклетки, пролиферацию гранулезных клеток, образование рецепторов ФСГ и ЛГ на поверхности гранулезных клеток, активность ароматаз в зреющем фолликуле (это усиливает конверсию андрогенов в эстрогены), продукцию ингиби-на, активина и инсулиноподобных факторов роста.
ЛГ способствует образованию андрогенов в текаклетках, обеспечивает овуляцию (совместно с ФСГ), стимулирует синтез прогестерона в лютеини-зированных клетках гранулезы (желтом теле) после овуляции.
Пролактин оказывает многообразное действие на организм женщины. Его основная биологическая роль - стимуляция роста молочных желез, регуляция лактации; он также обладает жиромобилизующим и гипотензивным эффектом, осуществляет контроль секреции прогестерона желтым телом путем активации образования в нем рецепторов к ЛГ. Во время беременности и лактации уровень пролактина в крови увеличивается. Ги-перпролактинемия приводит к нарушению роста и созревания фолликулов в яичнике (ановуляции).
Задняя доля гипофиза (нейрогипофиз) не является эндокринной железой, а лишь депонирует гормоны гипоталамуса (окситоцин и вазопрессин), которые находятся в организме в виде белкового комплекса.
Яичники относятся к четвертому уровню регуляции репродуктивной системы и выполняют две основные функции. В яичниках происходят циклические рост и созревание фолликулов, созревание яйцеклетки, т.е. осуществляются генеративная функция, а также синтез половых стероидов (эстрогенов, андрогенов, прогестерона) - гормональная функция.
Основной морфофункциональной единицей яичника является фолликул. При рождении в яичниках девочки находится примерно 2 млн приморди-альных фолликулов. Основная их масса (99%) в течение жизни подвергается атрезии (обратное развитие фолликулов). Только очень небольшая их часть (300-400) проходит полный цикл развития - от примордиального до преову-ляторного с образованием в последующем желтого тела. Ко времени менархе в яичниках содержится 200-400 тыс. примордиальных фолликулов.
Яичниковый цикл состоит из двух фаз: фолликулярной и лютеино-вой. Фолликулярная фаза начинается после менструации, связана с ростом
и созреванием фолликулов и оканчивается овуляцией. Лютеиновая фаза занимает промежуток после овуляции до начала менструации и связана с образованием, развитием и регрессом желтого тела, клетки которого се-кретируют прогестерон.
В зависимости от степени зрелости выделяют четыре типа фолликула: примордиальный, первичный (преантральный), вторичный (антральный) и зрелый (преовуляторный, доминантный) (рис. 2.2).

Рис. 2.2. Строение яичника (схема). Этапы развития доминантного фолликула и желтого тела: 1 - связка яичника; 2 - белочная оболочка; 3 - сосуды яичника (конечная ветвь яичниковой артерии и вены); 4 - примордиальный фолликул; 5 - преантральный фолликул; 6 - антральный фолликул; 7 - преовуляторный фолликул; 8 - овуляция; 9 - желтое тело; 10 - белое тело; 11 - яйцеклетка (ооцит); 12 - базальная мембрана; 13 - фолликулярная жидкость; 14 - яйценосный бугорок; 15 - тека-оболочка; 16 - блестящая оболочка; 17 - гранулезные клетки
Примордиальный фолликул состоит из незрелой яйцеклетки (ооцита) в профазе 2-го мейотического деления, которая окружена одним слоем гранулезных клеток.
В преантральном (первичном) фолликуле ооцит увеличивается в размерах. Клетки гранулезного эпителия пролиферируют и округляются, образуя зернистый слой фолликула. Из окружающей стромы формируется соедитель-нотканная оболочка - тека (theca).
Антральный (вторичный) фолликул характеризуется дальнейшим ростом: продолжается пролиферация клеток гранулезного слоя, которые продуцируют фолликулярную жидкость. Образующаяся жидкость оттесняет яйцеклетку к периферии, где клетки зернистого слоя образуют яйценосный бугорок (cumulus oophorus). Соединительнотканная оболочка фолликула отчетливо дифференцируется на наружную и внутреннюю. Внутренняя оболочка (the-ca interna) состоит из 2-4 слоев клеток. Наружная оболочка (theca externa) располагается над внутренней и представлена дифференцированной соединительнотканной стромой.
В преовуляторном (доминантном) фолликуле яйцеклетка, находящаяся на яйценосном бугорке, покрыта мембраной, называемой блестящей оболочкой (zona pellucida). В ооците доминантного фолликула возобновляется процесс мейоза. За время созревания в преовуляторном фолликуле происходит стократное увеличение объема фолликулярной жидкости (диаметр фолликула достигает 20 мм) (рис. 2.3).
В течение каждого менструального цикла от 3 до 30 примордиальных фолликулов начинают расти, преобразуясь в преантральные (первичные) фолликулы. В последующий менструальный цикл продолжается фоллику-логенез и только один фолликул развивается от преантрального до преову-ляторного. В процессе роста фолликула от преантрального до антрального

Рис. 2.3. Доминантный фолликул в яичнике. Лапароскопия
гранулезными клетками синтезируется антимюллеров гормон, способствующий его развитию. Остальные первоначально вступившие в рост фолликулы подвергаются атрезии (дегенерации).
Овуляция - разрыв преовуляторного (доминантного) фолликула и выход из него яйцеклетки в брюшную полость. Овуляция сопровождается кровотечением из разрушенных капилляров, окружающих текаклетки (рис. 2.4).
После выхода яйцеклетки в оставшуюся полость фолликула быстро врастают образующиеся капилляры. Гранулезные клетки подвергаются лютеи-низации, морфологически проявляющейся в увеличении их объема и образовании липидных включений - формируется желтое тело (рис. 2.5).

Рис. 2.4. Фолликул яичника после овуляции. Лапароскопия

Рис. 2.5. Желтое тело яичника. Лапароскопия
Желтое тело - транзиторное гормонально-активное образование, функционирующее в течение 14 дней независимо от общей продолжительности менструального цикла. Если беременность не наступила, желтое тело регрессирует, если же происходит оплодотворение, оно функционирует вплоть до образования плаценты (12-я неделя беременности).
Гормональная функция яичников
Рост, созревание фолликулов в яичниках и образование желтого тела сопровождаются продукцией половых гормонов как гранулезными клетками фолликула, так и клетками внутренней теки и в меньшей степени - внешней теки. К половым стероидным гормонам относятся эстрогены, прогестерон и андрогены. Исходным материалом для образования всех стероидных гормонов служит холестерин. До 90% стероидных гормонов находятся в связанном состоянии и только 10% несвязанных гормонов оказывают свой биологический эффект.
Эстрогены подразделяются на три фракции с различной активностью: эстрадиол, эстриол, эстрон. Эстрон - наименее активная фракция, выделяется яичниками в основном в период старения - в постменопаузе; наиболее активная фракция - эстрадиол, она значима в наступлении и сохранении беременности.
Количество половых гормонов меняется на протяжении менструального цикла. По мере роста фолликула увеличивается синтез всех половых гормонов, но преимущественно эстрогенов. В период после овуляции и до начала менструации в яичниках преимущественно синтезируется прогестерон, выделяемый клетками желтого тела.
Андрогены (андростендион и тестостерон) продуцируются текаклет-ками фолликула и межуточными клетками. Их уровень на протяжении менструального цикла не меняется. Попадая в клетки гранулезы, андро-гены активно подвергаются ароматизации, приводящей к их конверсии в эстрогены.
Помимо стероидных гормонов, яичники выделяют и другие биологически активные соединения: простагландины, окситоцин, вазопрессин, релаксин, эпидермальный фактор роста (ЭФР), инсулиноподобные факторы роста (ИПФР-1 и ИПФР-2). Полагают, что факторы роста способствуют пролиферации клеток гранулезы, росту и созреванию фолликула, селекции доминирующего фолликула.
В процессе овуляции определенную роль играют простагландины (F 2a и Е 2), а также содержащиеся в фолликулярной жидкости протеолитические ферменты, коллагеназа, окситоцин, релаксин.
Цикличность деятельности репродуктивной системы определяется принципами прямой и обратной связи, которая обеспечивается специфическими рецепторами к гормонам в каждом из звеньев. Прямая связь состоит в стимулирующем действии гипоталамуса на гипофиз и последующем образовании половых стероидов в яичнике. Обратная связь определяется влиянием повышенной концентрации половых стероидов на вышележащие уровни, блокируя их активность.
Во взаимодействии звеньев репродуктивной системы различают "длинную", "короткую" и "ультракороткую" петли. "Длинная" петля - воздействие через рецепторы гипоталамо-гипофизарной системы на выработку половых гормонов. "Короткая" петля определяет связь между гипофизом и гипоталамусом, "ультракороткая" - связь между гипоталамусом и нервными клетками, которые под действием электрических стимулов осуществляют локальную регуляцию с помощью нейротрансмиттеров, нейропепти-дов, нейромодуляторов.
Фолликулярная фаза
Пульсирующая секреция и выделение ГнРГ приводят к высвобождению ФСГ и ЛГ из передней доли гипофиза. ЛГ способствует синтезу андроге-нов текаклетками фолликула. ФСГ воздействует на яичники и приводит к росту фолликула и созреванию ооцита. Вместе с этим увеличивающийся уровень ФСГ стимулирует продукцию эстрогенов в клетках гранулезы путем ароматизации андрогенов, образовавшихся в текаклетках фолликула, а также способствует секреции ингибина и ИПФР-1-2. Перед овуляцией увеличивается количество рецепторов к ФСГ и ЛГ в клетках теки и гра-нулезы (рис. 2.6).
Овуляция происходит в середине менструального цикла, через 12-24 ч после достижения пика эстрадиола, вызывающего увеличение частоты и амплитуды секреции ГнРГ и резкий преовуляторный подъем секреции ЛГ по типу "положительной обратной связи". На этом фоне активизируются протеолитические ферменты - коллагеназа и плазмин, разрушающие коллаген стенки фолликула и таким образом уменьшающие ее прочность. Одновременно отмечаемое повышение концентрации простагландина F 2a , а также окситоцина индуцирует разрыв фолликула в результате стимуляции ими сокращения гладких мышц и выталкивания ооцита с яйценосным бугорком из полости фолликула. Разрыву фолликула способствует также повышение в нем концентрации простагландина Е 2 и релаксина, уменьшающих ригидность его стенок.
Лютеиновая фаза
После овуляции уровень ЛГ снижается по отношению к "овуляторному пику". Однако данное количество ЛГ стимулирует процесс лютеинизации гранулезных клеток, оставшихся в фолликуле, а также преимущественную секрецию образовавшимся желтым телом прогестерона. Максимальная секреция прогестерона происходит на 6-8-й день существования желтого тела, что соответствует 20-22-му дню менструального цикла. Постепенно, к 28-30-му дню менструального цикла уровень прогестерона, эстрогенов, ЛГ и ФСГ снижается, желтое тело регрессирует и заменяется соединительной тканью (белое тело).
Пятый уровень регуляции репродуктивной функции составляют чувствительные к колебаниям уровня половых стероидов органы-мишени: матка, маточные трубы, слизистая оболочка влагалища, а также молочные железы, волосяные фолликулы, кости, жировая ткань, ЦНС.
Стероидные гормоны яичников влияют на обменные процессы в органах и тканях, имеющих специфические рецепторы. Эти рецепторы могут быть

Рис. 2.6. Гормональная регуляция менструального цикла (схема): а - изменения уровня гормонов; б - изменения в яичнике; в - изменения в эндометрии
как цитоплазматическими, так и ядерными. Цитоплазматические рецепторы строгоспецифичны к эстрогенам, прогестерону и тестостерону. Стероиды проникают в клетки-мишени, связываясь со специфическими рецепторами - соответственно к эстрогенам, прогестерону, тестостерону. Образовавшийся комплекс поступает в ядро клетки, где, соединяясь с хроматином, обеспечивает синтез специфических тканевых белков через транскрипцию матричной РНК.
Матка состоит из наружного (серозного) покрова, миометрия и эндометрия. Эндометрий морфологически состоит из двух слоев: базального и функционального. Базальный слой в течение менструального цикла существенно не изменяется. Функциональный слой эндометрия претерпевает структурные и морфологические изменения, проявляющиеся последовательной сменой стадий пролиферации, секреции, десквамации с последующей
регенерацией. Циклическая секреция половых гормонов (эстрогены, прогестерон) приводит к двухфазным изменениям эндометрия, направленным на восприятие оплодотворенной яйцеклетки.
Циклические изменения в эндометрии касаются его функционального (поверхностного) слоя, состоящего из компактных эпителиальных клеток, которые отторгаются во время менструации. Базальный слой, не отторгаемый в этот период, обеспечивает восстановление функционального слоя.
В эндометрии в течение менструального цикла происходят следующие изменения: десквамация и отторжение функционального слоя, регенерация, фаза пролиферации и фаза секреции.
Трансформация эндометрия происходит под влиянием стероидных гормонов: фаза пролиферации - под преимущественным действием эстрогенов, фаза секреции - под влиянием прогестерона и эстрогенов.
Фаза пролиферации (соответствует фолликулярной фазе в яичниках) продолжается в среднем 12-14 дней, начиная с 5-го дня цикла. В этот период образуется новый поверхностный слой с вытянутыми трубчатыми железами, выстланными цилиндрическим эпителием с повышенной митотической активностью. Толщина функционального слоя эндометрия составляет 8 мм (рис. 2.7).
Фаза секреции (лютеиновая фаза в яичниках) связана с активностью желтого тела, длится 14±1 день. В этот период эпителий желез эндометрия начинает вырабатывать секрет, содержащий кислые гликозаминогликаны, гликопротеиды, гликоген (рис. 2.8).

Рис. 2.7. Эндометрий в фазе пролиферации (средняя стадия). Окраска гематоксилином и эозином, × 200. Фото О.В. Зайратьянца

Рис. 2.8. Эндометрий в фазе секреции (средняя стадия). Окраска гематоксилином и эозином, ×200. Фото О.В. Зайратьянца
Активность секреции становится наивысшей на 20-21-й день менструального цикла. К этому времени в эндометрии обнаруживается максимальное количество протеолитических ферментов, а в строме происходят децидуаль-ные превращения. Отмечается резкая васкуляризация стромы - спиральные артерии функционального слоя извиты, образуют "клубки", вены расширены. Такие изменения в эндометрии, отмечаемые на 20-22-й день (6-8-й день после овуляции) 28-дневного менструального цикла, обеспечивают наилучшие условия для имплантации оплодотворенной яйцеклетки.
К 24-27-му дню в связи с началом регресса желтого тела и снижением концентрации продуцируемого им прогестерона трофика эндометрия нарушается, постепенно в нем нарастают дегенеративные изменения. Из зернистых клеток стромы эндометрия выделяются гранулы, содержащие релаксин, подготавливающий менструальное отторжение слизистой оболочки. В поверхностных участках компактного слоя отмечаются лакунарные расширения капилляров и кровоизлияния в строму, что можно обнаружить за 1 сут до начала менструации.
Менструация включает десквамацию, отторжение и регенерацию функционального слоя эндометрия. В связи с регрессом желтого тела и резким снижением содержания половых стероидов в эндометрии нарастает гипоксия. Началу менструации способствует длительный спазм артерий, приводящий к стазу крови и образованию тромбов. Гипоксию тканей (тканевой ацидоз) усугубляют повышенная проницаемость эндотелия, ломкость стенок сосудов, многочисленные мелкие кровоизлияния и массивная лейко-
цитарная инфильтрация. Выделяемые из лейкоцитов лизосомальные проте-олитические ферменты усиливают расплавление тканевых элементов. Вслед за длительным спазмом сосудов наступает их паретическое расширение с усиленным притоком крови. При этом отмечаются рост гидростатического давления в микроциркуляторном русле и разрыв стенок сосудов, которые к этому времени в значительной степени утрачивают механическую прочность. На этом фоне и происходит активная десквамация некротизирован-ных участков функционального слоя эндометрия. К концу 1-х суток менструации отторгается 2 / 3 функционального слоя, а его полная десквамация обычно заканчивается на 3-й день менструального цикла.
Регенерация эндометрия начинается сразу после отторжения некроти-зированного функционального слоя. Основой для регенерации являются эпителиальные клетки стромы базального слоя. В физиологических условиях уже на 4-й день цикла вся раневая поверхность слизистой оболочки оказывается эпителизированной. Далее опять следуют циклические изменения эндометрия - фазы пролиферации и секреции.
Последовательные изменения на протяжении цикла в эндометрии - пролиферация, секреция и менструация - зависят не только от циклических колебаний уровня половых стероидов в крови, но и от состояния тканевых рецепторов к этим гормонам.
Концентрация ядерных рецепторов эстрадиола увеличивается до середины цикла, достигая пика к позднему периоду фазы пролиферации эндометрия. После овуляции наступает быстрое снижение концентрации ядерных рецепторов эстрадиола, продолжающееся до поздней секреторной фазы, когда их экспрессия становится значительно ниже, чем в начале цикла.
Функциональное состояние маточных труб варьирует в зависимости от фазы менструального цикла. Так, в лютеиновую фазу цикла активируются реснитчатый аппарат мерцательного эпителия и сократительная активность мышечного слоя, направленные на оптимальный транспорт половых гамет в полость матки.
Изменения в экстрагенитальных органах-мишенях
Все половые гормоны не только определяют функциональные изменения в самой репродуктивной системе, но и активно влияют на обменные процессы в других органах и тканях, имеющих рецепторы к половым стероидам.
В коже под влиянием эстрадиола и тестостерона активизируется синтез коллагена, что способствует поддержанию ее эластичности. Повышенная сальность, акне, фолликулиты, пористость кожи и избыточное оволосение возникают при повышении уровня андрогенов.
В костях эстрогены, прогестерон и андрогены поддерживают нормальное ремоделирование, предупреждая костную резорбцию. Баланс половых стероидов влияет на метаболизм и распределение жировой ткани в женском организме.
С воздействием половых гормонов на рецепторы в ЦНС и структурах гиппокампа связано изменение эмоциональной сферы и вегетативных
реакций у женщины в дни, предшествующие менструации, - феномен "менструальной волны". Этот феномен проявляется разбалансированием процессов активации и торможения в коре головного мозга, колебаниями симпатической и парасимпатической нервной системы (особенно влияющей на сердечно-сосудистую систему). Внешними проявлениями данных колебаний служат изменения настроения и раздражительность. У здоровых женщин эти изменения не выходят за пределы физиологических границ.
Влияние щитовидной железы и надпочечников на репродуктивную функцию
Щитовидная железа вырабатывает два йодаминокислотных гормона - трийодтиронин (Т 3) и тироксин (Т 4), которые являются важнейшими регуляторами метаболизма, развития и дифференцировки всех тканей организма, особенно тироксин. Гормоны щитовидной железы оказывают определенное влияние на белково-синтетическую функцию печени, стимулируя образование глобулина, связывающего половые стероиды. Это отражается на балансе свободных (активных) и связанных стероидов яичника (эстрогенов, андрогенов).
При недостатке Т 3 и Т 4 повышается секреция тиреолиберина, активизирующего не только тиреотрофы, но и лактотрофы гипофиза, что нередко становится причиной гиперпролактинемии. Параллельно снижается секреция ЛГ и ФСГ с торможением фолликуло- и стероидогенеза в яичниках.
Возрастание уровня Т 3 и Т 4 сопровождается значительным увеличением концентрации глобулина, связывающего половые гормоны в печени и приводящего к уменьшению свободной фракции эстрогенов. Гипоэстрогения, в свою очередь, приводит к нарушению созревания фолликулов.
Надпочечники. В норме продукция андрогенов - андростендиона и тестостерона - в надпочечниках оказывается такой же, как и в яичниках. В надпочечниках происходит образование ДГЭА и ДГЭА-С, тогда как в яичниках эти андрогены практически не синтезируются. ДГЭА-С, се-кретируемый в наибольшем (по сравнению с другими надпочечниковыми андрогенами) количестве, обладает относительно невысокой андрогенной активностью и служит своего рода резервной формой андрогенов. Надпо-чечниковые андрогены наряду с андрогенами яичникового происхождения являются субстратом для внегонадной продукции эстрогенов.
Оценка состояния репродуктивной системы по данным тестов функциональной диагностики
Многие годы в гинекологической практике используются так называемые тесты функциональной диагностики состояния репродуктивной системы. Ценность этих достаточно простых исследований сохранилась до настоящего времени. Наиболее часто используют измерение базальной температуры, оценку феномена "зрачка" и состояние шеечной слизи (ее кристаллизация, растяжимость), а также подсчет кариопикнотического индекса (КПИ, %) эпителия влагалища (рис. 2.9).

Рис. 2.9. Тесты функциональной диагностики при двухфазном менструальном цикле
Тест базальной температуры основан на способности прогестерона (в возросшей концентрации) непосредственно воздействовать на центр терморегуляции в гипоталамусе. Под влиянием прогестерона во 2-ю (лютеи-новую) фазу менструального цикла происходит транзиторная гипертермическая реакция.
Пациентка ежедневно измеряет температуру в прямой кишке утром, не вставая с постели. Результаты отображают графически. При нормальном двухфазном менструальном цикле базальная температура в 1-ю (фолликулярную) фазу менструального цикла не превышает 37 °C, во 2-ю (лютеиновую) фазу отмечается повышение ректальной температуры на 0,4-0,8 °C по сравнению с исходным значением. В день менструации или за 1 сут до ее начала желтое тело в яичнике регрессирует, уровень прогестерона уменьшается, в связи с чем базальная температура снижается до исходных значений.
Стойкий двухфазный цикл (базальную температуру нужно измерять на протяжении 2-3 менструальных циклов) свидетельствует о произошедшей овуляции и функциональной полноценности желтого тела. Отсутствие подъема температуры во 2-ю фазу цикла указывает на отсутствие овуляции (ановуляцию); запаздывание подъема, его кратковременность (повышение температуры на 2-7 дней) или недостаточный подъем (на 0,2-0,3 °C) - на неполноценную функцию желтого тела, т.е. недостаточность выработки прогестерона. Ложноположительный результат (повышение базальной температуры при отсутствии желтого тела) возможен при острых и хронических инфекциях, при некоторых изменениях ЦНС, сопровождающихся повышенной возбудимостью.
Симптом "зрачка" отражает количество и состояние слизистого секрета в канале шейки матки, которые зависят от эстрогенной насыщенности организма. Феномен "зрачка" основан на расширении наружного зева церви-кального канала из-за накопления в нем прозрачной стекловидной слизи и оценивается при осмотре шейки матки с помощью влагалищных зеркал. В зависимости от выраженности симптом "зрачка" оценивается по трем степеням: +, ++, +++.
Синтез цервикальной слизи в течение 1-й фазы менструального цикла увеличивается и становится максимальным непосредственно перед овуляцией, что связано с прогрессирующим увеличением уровня эстрогенов в данный период. В преовуляторные дни расширенное наружное отверстие канала шейки матки напоминает зрачок (+++). Во 2-ю фазу менструального цикла количество эстрогенов уменьшается, в яичниках преимущественно вырабатывается прогестерон, поэтому количество слизи уменьшается (+), а перед менструацией и вовсе отсутствует (-). Тест нельзя использовать при патологических изменениях шейки матки.
Симптом кристаллизации цервикальной слизи (феномен "папоротника") При высушивании максимально выражен во время овуляции, затем кристаллизация постепенно уменьшается, а перед менструацией вообще отсутствует. Кристаллизация слизи, высушенной на воздухе, оценивается также в баллах (от 1 до 3).
Симптом натяжения шеечной слизи прямо пропорционален уровню эстрогенов в женском организме. Для проведения пробы корнцангом извлекают слизь из шеечного канала, бранши инструмента медленно раздвигают, определяя степень натяжения (расстояния, на котором слизь "разрывается"). Максимальное растяжение цервикальной слизи (до 10-12 см) происходит в период наибольшей концентрации эстрогенов - в середине менструального цикла, что соответствует овуляции.
На слизь могут негативно влиять воспалительные процессы в половых органах, а также нарушения гормонального баланса.
Кариопикнотический индекс (КПИ). Под влиянием эстрогенов происходит пролиферация клеток базального слоя многослойного плоского эпителия влагалища, в связи с чем в поверхностном слое увеличивается количество ороговевающих (отшелушивающихся, отмирающих) клеток. Первой стадией гибели клеток являются изменения их ядра (кариопикноз). КПИ - это отношение числа клеток с пикнотическим ядром (т.е. ороговевающих) к общему числу эпителиальных клеток в мазке, выраженное в процентах. В начале фолликулярной фазы менструального цикла КПИ составляет 20- 40%, в преовуляторные дни повышается до 80-88%, что связано с прогрессирующим увеличением уровня эстрогенов. В лютеиновой фазе цикла уровень эстрогенов снижается, следовательно, КПИ уменьшается до 20- 25%. Таким образом, количественные соотношения клеточных элементов в мазках слизистой оболочки влагалища позволяют судить о насыщенности организма эстрогенами.
В настоящее время, особенно в программе экстракорпорального оплодотворения (ЭКО), созревание фолликула, овуляция и образование желтого тела определяются при динамическом УЗИ.
Контрольные вопросы
1. Охарактеризуйте нормальный менструальный цикл.
2. Укажите уровни регуляции менструального цикла.
3. Перечислите принципы прямой и обратной связи.
4. Какие изменения происходят в яичниках в течение нормального менструального цикла?
5. Какие изменения происходят в матке в течение нормального менструального цикла?
6. Назовите тесты функциональной диагностики.
Гинекология: учебник / Б. И. Баисова и др. ; под ред. Г. М. Савельевой, В. Г. Бреусенко. - 4-е изд., перераб. и доп. - 2011. - 432 с. : ил.
| Статьи по теме: | |
|
Журнал осмотра детей на чесотку и педикулез — форма и оформление Журнал осмотра контактных детей
Отрасль: Образование Специализации: Заведующему детским садом,... Это (,) наверное (,) вводные слова
Выражение «на самом деле» запятой может отделяться от остального... Несколько способов гадания на свадьбу: все карты Вам в руки
Для любой девушки очень волнительным и важным вопросом считается... | |




